上帝的基督教会
[No. 265]
万民的遗传根源
(版本4.0 20060105-20060421-20070804-20071122)
圣经清楚及明确地叙述了万民的出生,这被指与现代的科学主张不一致。当我们以历史证据和已知的族群迁移对证科学记录时,我们发现圣经与科学可以协同一致。问题在于进化论者基于错误假设所建立的,被大大地拉长的数学模型。
Christian Churches of God
PO Box 369, WODEN ACT 2606, AUSTRALIA
Email: secretary@ccg.org
(版权© 2006, 2007 Wade Cox)
本文只可在未经任何删改的情形下允许自由复制和散发。出版者的名字和地址以及版权通告必须包含在内。不可对收得散发副本者索取收费。在评论性文章和论述中所作的简短引述不侵犯版权。
从互联网页可取得此文:
http://www.logon.org和 http://www.ccg.org
万民的遗传根源
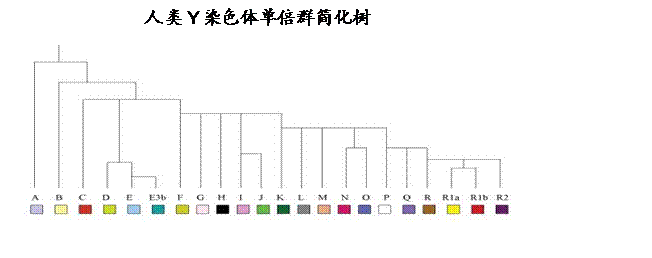
人类遗传学的最大进展之一是对男性和女性脱氧核糖核酸(DNA)的测定。男性是通过连续世系从父传给子,随时间突变的Y染色体来测定。这称为人类Y染色体(YDNA)。只有男性,作为XY,通过他们的Y染色体相传。女性有XX染色体。科学说这突变的速度比圣经要求符合万民系出诺亚的历史慢得多。
母系是通过她们的线粒体脱氧核糖核酸(称mtDNA) 检测,男性和女性都有线粒体脱氧核糖核酸。线粒体是在它们所出现,与母系有关的单倍群(Haplogroup)组内,这系从母亲在X染色体中传给子女;男性和女性都有X染色体。
人类Y染色体和线粒体脱氧核糖核酸是以两个不同的方式检测。人类Y染色体是以所谓的多形体来检测。这些多形体被分配一个数值,在检验时根据这数值,形成的子分组被称为整体分子即单倍群的分化体和子分化体。这些分值记录人类Y染色体突变和系的变化。
线粒体脱氧核糖核酸是通过报告多态性位点例如311C来检定,意思是碱基对发生了突变。这里16,311和变化的碱实际上变成了胞嘧啶。16,000是脱氧核糖核酸编码的开始点,因此16,000被省略而被使用的编码是超逾的数字。因此16,311成为311,而有关的字母指示多形体中那一点的化学物。多态性位点的改变即是决定遗传世系者,因为父母将他们所有的相同或类似数值脱氧核糖核酸多态性传给子女。在检测时,这些与父母不完全相同的数值被称为突变。其等级因此不同并决定了世界万民的部族分组。
配给男性人种的人类Y染色体系统从A至R被分组成一系列的单倍群。一向的广泛检测(使用亚利桑那系统)通常是以三十七位点为遗传标记。 前十二是基本检测,然后是二十五,然后继续至这些多态性位点,或位置的三十七点,以测定关联性和单倍群关联。可以有大约一百或更多遗传标记的变化(又名多态性)检测。人类Y染色体典型结构的可能例子如下:
1 |
393 |
x |
2 |
390 |
x |
3 |
19* |
x |
4 |
391 |
x |
5 |
385a |
x |
6 |
385b |
x |
7 |
426 |
x |
8 |
388 |
x |
9 |
439 |
x |
10 |
389-1 |
x |
11 |
392 |
x |
12 |
389-2 |
x |
13 |
458 |
x |
14 |
459a |
x |
15 |
459b |
x |
16 |
455 |
x |
17 |
454 |
x |
18 |
447 |
x |
19 |
437 |
x |
20 |
448 |
x |
21 |
449 |
x |
22 |
464a** |
x |
23 |
464b** |
x |
24 |
464c** |
x |
25 |
464d** |
x |
26 |
460 |
x |
27 |
GATA H4 |
x |
28 |
YCA II a |
x |
29 |
YCA II b |
x |
30 |
456 |
x |
31 |
607 |
x |
32 |
576 |
x |
33 |
570 |
x |
34 |
CDY a |
x |
35 |
CDY b |
x |
36 |
442 |
x |
37 |
438 |
x |
*也被称为 DYS 394
**在2003年五月十九日,这些数值因实验室命名的一项改变而被调下一点。
有些数值在检测后被配至表的右栏。这些数值协助检定一个人是否落入一个 典型的凯尔特单倍群或在闪米特人,或印第安人,或波利尼西亚人,或华人或日本人种族类型中所发现的单倍群。要适当地鉴定这类型,得进行一项称为单核苷酸多态性(SNP)的检测 ,但从12, 25和37标记检测所得数值将我们指向正确的方向。没有一个民族群体属纯遗传单倍群,除了南非Q分组的一些印第安人部族之外(从J. D. 麦克唐纳医生制作的图表)。以上的表,视检测数值而定,可能属于欧洲的典型R1b 凯尔特或美洲原住民,他们的脱氧核糖核酸皆属于表中的最后两个单倍群。
线粒体脱氧核糖核酸
线粒体脱氧核糖核酸,于1981年首次被排序,被称为剑桥参考序列(CRS)。这系列被用作与单一线粒体脱氧核糖核酸作比较的基础。换言之,单一线粒体脱氧核糖核酸中任何与剑桥参考序列有差异的位置将被称为突变。 若结果显示全无突变,则表示那线粒体脱氧核糖核酸与剑桥参考序列相称。当突变发生时:a) 一碱基取代另一碱基-例如胞嘧啶(C) 取代了腺嘌呤(A);b)一碱基不再在那方位,或置,或缺失;以及c)一个新碱基在未取代任何其他碱基的情况下插入其他碱基之间(掺入)。
这是线粒体脱氧核糖核酸突变的情况。人种的原有女性脱氧核糖核酸只局限于七女性,但只在欧洲而已。全世界有大约廿六个母系人类树子分支。因此,母系可能可争议为只有欧洲可在圣经术语中解释。因此须解释全部廿六个母系使线粒体脱氧核糖核酸与圣经的叙述 一致。
我们依赖科学对单倍群的组织。在多形体突变时它们的数值改变因而突变率可能导致仍未被确定的单倍群之间的一些运动。
使圣经和遗传学协同一致
若圣经要答覆这非常重要遗传发现所构成的挑战,它必须将这结构置于圣经记录之上以及在可考虑到、并正确回答人类Y染色体和这些种类的线粒体脱氧核糖核酸两者的遗传树所构成挑战的说法之内解释万民的分布。
母系可能会对男性产生影响,导致仍未知的突变,对那系统的研究可能也将揭示一些重要的层面。无论如何,目前除了所知的基因重组,未有科学证明女性对男性基因突变有直接影响。
然而,我们现在知道线粒体是直接造成人类遗传结构突变的原因。它掺入活性基因并导致突变。这特别是在被污染或辐射损伤的脱氧核糖核酸方面,使到它们易于受到流离线粒体脱氧核糖核酸的进一步损伤。操作基因较易于受到损伤,因为它们的脱氧核糖核酸被定期解开以生产蛋白(新科学家, 2004年九月十八日,文章-"那靶向基因内的敌人",页11)。
“巴黎巴斯德研究院的米里亚里凯蒂和她的工作队确定了横跨基因组的二百一十一个线粒体脱氧核糖核酸掺入,其中的二十三个是新的,80%是在基因中但基因组成基因组的3%。里凯蒂说,靶向基因是广泛地被转录的基因,因为它们有较多的双链断裂。”(如上)。
约翰霍普金斯大学的耶夫博克认为线粒体脱氧核糖核酸不大可能是人类基因突变的主要原因。不过,这似有一直接关系到切尔诺贝利核电站灾祸的案例。
通过比较二十一名不同种族志愿者的序列,里凯蒂的工作队发现六个不为所有人享有的掺入(普洛斯生物学期刊,册2, e273)。这些科学家认为“这些必是在我们的祖先从猩猩的祖先分裂后发生因此可为科学家提供关于族群的来源和历史迁移的线索。”(如上)
我们同意这些线粒体有能力使人类遗传结构产生突变,而与新线粒体异族通婚的发生率被认为是导致突变加快速度的原因。
那是为何孤立社会比与其他线粒体单倍群混合的开放社会显示着基本结构和较小的改变。例如,澳洲原住民之中的RxR1组也受较小影响。它非时间指标,而是异族通婚。不过,原住民单倍群C的大部分实际上是C4,并衍生自散开在印度至越南南亚海岸一带的C基组。 有一些C基仍在襄里,但原住民在澳洲内发展成为C4组然后再分至两个C4子群。毛利人发展成为C2组而蒙古族成为 C3,它也进入美国。原住民和毛利语言两者都包含很多借自马来人的词表示他们曾经分享一个共同地区和语言群体。不过,马来人主要是单倍群O但也有一些其他类群。毛利人在水位升涨后约于公元1000年在中世纪暖期当海域较高期间离开他们的岛屿家园前往纽西兰。
称为Y染色体种系树似有个问题,因单倍群A是这树第一个显著的分叉,因而是第一个从单倍群B分歧至R者。所有在分歧处都有一个叉。
现在我们来讨论一个圣经记述概述。圣经简单地说诺亚在他的时代是完美的,而他生了雅弗、然后含接着闪。诺亚众子的后代被列出而且族群和分布也被记载,因此给予我们一些宝贵的线索来与脱氧核糖核酸记录作对照。
圣经的记述被解释为只允许八个人在方舟上。不过,这记录可能指的是八个男性,因为除了非常罕见的例子以及有时只在删除父亲姓名中的前缀之外,女性在古代系谱中从未被提及(见文章弥赛亚的系谱(No. 119))。
这八男情况是很可能的,因迦南在洪水过后第一个葡萄收获中,当诺亚喝新酒醉了或鸡奸他或阉割他时年龄已够大了。犹太传说讨论这两种情景。这是圣经诅咒迦南的原因。一些指它是含,而迦南承受这诅咒而非含,但那似乎是要局限这八口于男性和女性。后一种情况对脱氧核糖核酸记录是重要的,在于只有儿子的妻子会继续繁殖而诺亚妻子的线粒体脱氧核糖核酸将已停止,除非也有来自诺亚妻子的女性在那里,若只有男性被提及那是可能的。
父系因此是雅弗和闪与他们的妻子,以及含和含的(四个)儿子和他们的妻子,总共是七男七女,还有诺亚和他的妻子。
这也将允许八个女性,包括诺亚的妻子,因此提供主要的线粒体脱氧核糖核酸母系;而那似与线粒体脱氧核糖核酸单倍群的现有发现一致,但仅限于欧洲。或可争论一些线粒体脱氧核糖核酸世系是后期的分组。科学证据显示高加索人中的线粒体脱氧核糖核酸基因库中有七个所谓的“夏娃”但总共有廿六个母系。因此,有关的解释必须说明整个序列。因此可提出方舟上共有廿六名女性继续繁殖的论点。
就母系而言,这观点将使科学与圣经记录直接协调一致。我们也将发现,当我们探讨母系 线粒体脱氧核糖核酸的记录时,我们将能够使女性分组与人类Y染色体男性分组取得一致。因此我们能够看到语亚的家庭在他们世界分布中的分解。我们较后将探讨这问题。
诺亚基因库
诺亚被理解为在他的世代是纯洁的。圣经也强调方舟里的人都是诺亚的家属。因此,要适当地说明遗传的多样性,诺亚必须保持丢弃基因独特后代的能力,而这后代拥有它所来自世系的特征,但非诺亚原本拥有的的整个序列。
人类Y染色体系统的传统观点是,它们回数 – 三万五千年,一些说六万年至十三万–以及突变的水平缓慢。这扩展的观点是基于数学模型但似乎是为提倡一个进化模型。此外,多元的女性脱氧核糖核酸系统的影响被置之不理。线粒体脱氧核糖核酸被检测,但女性X染色体对男性Y染色体结构的影响被认为是重要的,而且它会被争论说它使到人类Y染色体结构的突变比现有模型相信可允许的速度要快得多。
我们能够研究的其中一个已知历史是犹太人的历史而且我们可以鉴定一些出自他们的民族;在他们的脱氧核糖核酸中我们可看到很多人根据现有智慧实际上并不是闪米特族。例如,根据人类Y染色体模型和现有解释,所有阿什肯纳兹人的其中52%并非闪米特族。因此,我们在此可能有证据显示有众多皈依犹太教者与这族人结婚。它很可能指向可萨人的皈依为阿什肯纳兹利未人的来源。我们将在下面探讨这方面。
此外,我们知道津巴布韦连巴人的布巴族在Y染色体世系中都是利未的柯希宁族,而连巴人主要是犹太Y染色体,虽然是黑的;他们在巴比伦被掳时被隔离或之后娶了非洲妻子。他们的犹太教比被掳后的犹太教更基本而且他们的日历决定法是基于提前确定的日月会合。他们与犹大分开后以非常基本的形式坚持他们的信仰达二千五百年。
本文中的所有脱氧核糖核酸分布区是根据约1500年欧洲分散前根据检测的已知土著人口。
诺亚作为人类结构之父,他被认为得有以下人类Y染色体亚结构的能力,因为所有人类都是他的后裔。地球上的任何男性只会有象征他的分支和迹线的突变。
M91, SRY 10831a, M42, M94, M139, M60, M181, RPS4Y711, M216, YAP M145, M203, M174, SRY 4064, M96, P29, P14, M89, M213 和随附异数的核心。
诺亚拥有能够突变成这些其他子群的基本人类Y染色体。从源于他的分组如下:
含的儿子:
人类Y染色体单倍群A: M91 和附随异数
A1: M31
A2: M6, M14, M33, M49, M71, M135, M141, P3, P4, P5, MEH1, M196, M206
A3: M32
A3a: M28, M59
A3b M144, M190, M220
A3b1: M51
A3b2: M13, M63, M127
A3b2a: M171
A3b2b: M118
这树与其他单倍群B 至R不同。
这一组主要被发现在苏丹>45%, 埃塞俄比亚> 15%, 和南非> 30%。非常小的单倍群A元素被发现在喀麦隆,表面上在摩洛哥,以及中非的俾格米人之中。这些人的大部份是俾格米人和南非是ExE3b 和B,或摩洛哥E3b。
非洲主要是ExE3b 和E3b,有B组分布在他们之中。在非洲发现的A,在留在非洲的一组和离开非洲的一个支部之前,也有单倍群C为其根源。
单倍群B 是由M60, M181组成,然后:
B1: M146
B2: M182
B2a: M150
B2a1: M109, M152, P32
B2a2: M108a
B2a2a: M43
B2b1: P6
B2b2: M115, M169
B2b3: M30, M129
B2b3a: M108b
B2b4: P7
B2b4a: P8
B2b4b: MSY2a, M211
单倍群B 在马里、喀麦隆、中非俾米人之中、在南非、在苏丹和在埃塞俄比亚出现的百分比较小。单倍群A和B在非洲或非裔美国人之外的出现微不足道。
单倍群C
单倍群C是非常有趣的单倍群。它衍生自脱氧核糖核酸M168和P9组,然后至 成为基本C组的RPS4Y711和M216分节。基本C组在澳洲原住民中发现。其分支是:
C1: M8, M105, M131
C2: M38
C2a: P33
C3: M217, P44
C3a: M93
C3b: P39
C3c: M48, M77, M86
C4a
C4b
C组在以下民族中发现:
澳洲原住民>65% (C4)
毛利人>80% (C2)
法属玻里尼西亚人>55%
西萨摩亚人 >30%
伊里安加亚,婆罗洲(加里曼丹), 新不列颠及菲律宾发现较小的百分比。
中国和日本也发现较小的百分比。
主要的百分比发现在北亚:
蒙古族高达60%
布里亚特族高达65%
鄂温克族高达70%
科里亚克族高达35%
阿尔泰山族高达23%
哈萨克族高达42%
乌兹别克族高达15%
柯尔克孜族高达10%
雅库特族高达10%
有一元素(C3)移入阿拉斯加的印第安人(说约42%),然后至加拿大的齐帕威或纳得内部族 (+- 6%)以及美国的晒延人(>14%)和阿帕切人(>12%)。有很小百分比的格陵兰爱斯基摩人显示广泛的北极接触。
我们能够掘出亚洲这些人和部族的起点是具有重大意义的。中国的周口店上层洞穴(于1930年发现并由裴文中所挖掘1938-1940) 被发现是居住在同一层一个家庭组的家园,由美拉尼西亚、爱斯基摩和蒙古族类组成。魏敦瑞指瓦贾克遗骸较像周口店上层洞穴的成年女性遗骸,至今比任何地中海人更像(R.M.和C.H. 伯恩特,澳洲土著,安格斯与罗柏逊出版社,1965, 页30, 32-33)。
赫克斯利(1870)过去曾把澳洲土著分级为德拉维亚人的前卫;他们离开地中海和埃及进入印度然后到澳洲。他视德坎山族,以及更无关紧要地,古代埃及人,为与澳洲土著的唯一联系(如上,页33)。他将早期的人类分为四种:黑人、蒙古人、金发白种人和澳大利亚土著。塔斯马尼亚的土著他分级为黑人的美拉尼西亚分支。华莱士(1893年)曾说,澳洲人黑人也不是蒙古族,并将他们分级为早期的高加索人(如上)。不过,这已被显示为不正确的,而且澳洲土著是离开非洲或中东的单倍群C分支的直接后裔。他们也包括另两个单倍群,即美拉尼西亚人的单倍群 K和RxR1基,它实际上是一个基本亚利安单倍群,R1a斯拉夫人和R1b欧洲人是其后代。
现在我们从脱氧核糖核酸知道在周口店洞穴的发现具这三类型的同一家庭事实上是正确的。 这些族群的Y染色体显示这些人的确全传自相同的父系,而澳洲人是取道北部从非洲进入澳洲的基本C结构;美拉尼西亚人和波利尼西亚人发展太平洋,而蒙古人以及成为远北族群者则进入了北亚。
据作者所知并无联系现有澳洲土著与较早在澳洲的发现之早期澳大利亚土著脱氧核糖核酸的公布。澳洲有两个主要人类Y染色体组,它们是C4 和K 单倍群,有早期RxR1 单倍群>10%,只在非洲的喀麦隆(>40%)有重要的发现,在乌兹别克斯坦和德拉维人之中有较小的组,在任何情况下都很少见。那RxR1组将证明与非洲和印度德拉维人的联系。不过,那是澳洲土著与德拉维人的唯一联系。C4组是较好的联系,因此大体上土著来自形成非洲组的群组,而那也形成喀麦隆和东北非的群组。
大部分的欧洲人是 R1a 和R1b,而且可能被视为传自这较早的RxR1 脱氧核糖核酸联系。无论如何,RxR1单倍群被认为是排除这些土著的任何古代起源的。K单倍群的25%发生率显示早期(最初)美拉尼西亚人从北部进入澳洲的迁移。C组似来自非洲/中东进入中亚。这方面可从圣经解释为古实人的分裂。西方古实人通过苏丹进入非洲,而东方古实人先进入印度然后分组继续移动。在亚利安人于公元前侵略印度后,他们似搬离了印度。这些土著在南亚海岸与C基组分离,可能因他们的游牧存在,并可能利用独木舟南行进入澳洲。他者东行和北行进入亚洲,然后太平洋。因此,非洲来源或可用在中东的分裂而非从亚洲的迁移来作解释,因而不但是这些土著的元素,C组的整个蒙古和太平洋系统有着中亚来源。R 基组被争论为在印度/巴基斯坦形成而并未指示随迁移进入非洲和澳洲的中东来源。我们将在处理单倍群R时讨论这问题。这些土著的多元化语言群体(八)显示至少在三个部族单倍群系统方面有大约八个进入澳洲的浪潮或可能有更多不同的移民。
南美洲没有C 单倍群,因此毛利人和南美洲人之间的假设联系是个神话。美国的主要人类Y染色体是 Q 单倍群,C数量少得多,只在从阿拉斯加进入北美洲的纳得内人、晒延人和阿帕切人以及中美的玛雅人之中。
C组被认为可能在图表中被错误配置而且或许应与形成单倍群K和M的原始美拉尼西亚人的K组有较密切的联系。 它也许可能与单倍群E换位,但在以上A和C论点的情况下那可能不是这样的。C组的迁移是与进入美国Q组一起的次要群体。一般看法指他们在一万二千至一万五千年前迁移,但圣经的时帧不是这么说。
毛利人对作者说,叙述这族群迁移的方舟传说以及将他们的世系联系于那些抵达纽西兰方舟的传说,实际上允许毛利人有非洲根源。然而,C组脱氧核糖核酸随着O情况的出现,似要求中亚根源而从中国海岸的迁移或与汉人的扩展相符。 无论如何,他们的马来语言联系显示南迁进入印度马来群岛,毛利人和土著在此通过隔离分裂为C4和C2。毛利人中有东和东南亚 (O)Y染色体踪迹,但在西萨摩亚人中有更大的元素,以及在稍少的程度上,在法属玻里尼西亚人之中。这可能显示马来/菲律宾人对C系统太平洋族群的影响,但这种影响在我们预期更容易找到它的伊里安查亚、巴布亚新几内亚,和新布列颠中并不显著。毛利人相当迟抵达纽西兰,他们在现有纪元的第二千年早期世纪中取代了纽西兰过去的两个文化。第一个是被迫迁移的巴布亚型文化,后者在它的石结构中似近乎是凯尔特人。从C和RxR1组较早期澳洲土著的迁移看来,他们的迁移因此相当明显。
单倍群D
这单倍群是在YAP分支从D和E组形成,M145和M203是两者的母体分成D:
D1: M15
D2: M55, M57, M64a, P37a, P41a, 12f2b
D2a: P12, P42
D2b: M116a
D2B1: M125
D2b2: M151
这单倍群在任何意这义上局限于两种人。他们是日本人(>40%)和西藏人(>50%)。单倍群的出现显示日本人和西藏人曾经是占据东方干草原的民族,他们向东北迁移进入日本向南移进入西藏山脉。日本的语言是乌拉尔阿尔泰语,与芬兰和匈牙利、土耳基、阿尔泰、蒙古、满族/通古斯、老韩和北西伯利亚语言(以及在古代与巴斯克语)有很多共同之处。因此,我们可以假设单倍群D与 单倍群C 和以下鉴定的他者有一个共同的语言系统。日本人和西藏人的其他单倍群(O)显示中国人多个世纪来与他们异种繁殖,无疑影响了他们迁移地区的语言系统和风俗。日本人中单倍群C出现的比率比西藏人高,或是基于他们与蒙古人、布里亚特人和科里亚克族的接触。不过,由于后期日本侵略的人口置换,阿依努土著的情况也可以是它的因素之一。
其他显著的D发生率或来自日本早期前往苏门答腊和马来亚贸易的影响。较显著的出现是在苏门答腊,高达10%。马来亚少过5%,那里的R1a组也一样,而在F, C和M方面, K的出现率稍高。婆罗洲也有少过5%的D组,维吾尔、阿尔泰、蒙古、吉尔吉斯、乌兹别克和西伯利亚爱斯基摩人也一样。
单倍群E
单倍群E基本上在非洲发现。它在在M145 和M203继续Yap分支至SRY 4064 M96和P29,形成E基组。
E1: M33, M132
E1a: M44
E2: M75
E2a: M41
E2b: M54, M85, M90
E3: P2, DYS 391p
E3a: M2, P1
E3a1: M58
E3a2: M116b
E3a3: M149
E3a4: M154
E3a5: M165
E3a6: M10, M66, M156
E3b: M35, M215 (在一些图表中被省略但包含在脱氧核糖核酸2005 Y染色种系树中,见www.familytreeDNA.com)。
E3b1: M78
E3b1a: M148
E3b2: M81
E3b2a: M107
E3b2b: M165
E3b3: M123
E3b3a: M34
E3b3a1: M136
E组在非洲被发现,中东和南欧有较小的发现。一些人推测英国E3a的发现是源自布列颠的罗马奴隶;其他人视他们为后期到来的奴隶。两个观点都可能正确。
ExE3b在布基纳法索中最高,远超90%或达99%。
ExE3b在马里高达(60%), 喀麦隆(45%), 俾格米(65%)和南非(约55%)。这一组在苏丹和埃塞俄比亚中的出现率低。
E3b在摩洛哥很高(约75%),单倍群J的发现约达10%。马里有显著的发现(25%), 苏丹(25%), 以及埃塞俄比亚(55%,) ,南非(少过10%)。15%的中东阿拉伯人和波斯人是E3b, 约有10%是意大利人和伊比利亚人,5% 到10% 的德国人和苏联人是E3b。有达5%的格鲁吉亚人和亚美尼亚人是E3b。东欧的犹太人有25%是E3b。他们可能已从苏丹和埃塞俄比亚皈依犹太教,因两个地区都有闪米特元素。他们可能是出埃及中混合众的一部分。较可能的是,很多得幸免并在约书亚之下的占据后加入以色列。我们将在以下探讨闪米特单倍群 时讨论这一面。
这些单倍群A至E是含的儿子们。我们较后将设法鉴定他们的族群。
如所说的,单倍群C可能是被错误配置而应置于树里的E之后,F的旁边以及称为YAP分离的这一边。D和E是由D和E共有的,对Y的掺入为定义,而 C和F 没有这掺入。它们在部落地点较常出现在一起。
单倍群F
F 单倍群出自Yap M145, M203分至D和E以及RPS4Y 711 M216分至C组之间的分支。它是一个小组,有时充当一个杂库,因为研究者未作出足够的检测来正确地鉴定这一组。 格鲁吉亚/亚美尼亚、波斯、乌兹别克斯坦、喀山鞑靼之中有小F组。结论是,F的基本根将近消失但多产的子孙们幸免于难并繁荣起来,生产了主要的族群和来自这母体的突变。
这母体决定了F至 R的所有其他单倍群。
基本F是sP14, M89, M213。
从圣经而言,闪和雅弗将这核心单倍群传给他们所有的后代。或许含也传给他的儿子之一。有关的图表将使含的众子广泛分岔。
线粒体脱氧核糖核酸之谜
在我们进一步研究人类Y染色体单倍群之前,我们将提出一问题并建议一项方案。
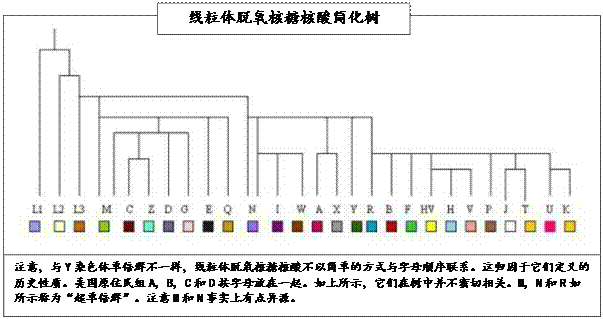
共有廿六个线粒体脱氧核糖核酸单倍群显示廿六个线粒体脱氧核糖核酸母系。有大约七个摆造给欧洲的固有单倍群或女“夏娃”。不过,当我们研究线粒体脱氧核糖核酸树时,我们发现一些有趣的群体派生物。这些所谓的“超群”事实上只在三个基组中。换句话说,它们来自三个主要的母系。若我们假设方舟上只有三名女性,即闪、含和雅弗的妻子继续繁殖,那将是我们预期会发现的。这些单倍群都出自一个单一的女性超群,即单倍群L。因此,事实上,所有女性都出自一个母系,单倍群L。那是超级L。这系然后分裂成L1,然后L2和L3。L3趋异而从L3产生其他线粒体脱氧核糖核酸突变。因此,所有女性出自一个夏娃,她的线粒体脱氧核糖核酸系是L。
L组L1, L2和L3都在亚洲发现,而且几乎是非洲撒哈拉沙漠以南地区独有的主要组群。只有从埃塞俄比亚北部我们可取得很多线粒体脱氧核糖核酸多样性的记录。那是进化论者宣称我们全都是出自非洲的基本原因。脱氧核糖核酸组的分置可在J. D.麦克唐纳的作品中看到;他以Y和线粒体脱氧核糖核酸单倍群图表将它们分组:
http://www.scs.uiuc.edu/~mcdonald/WorldhaplogroupsMaps.pdf.
超群M和N是下一个趋异或突变者。从圣经观点,我们可轻易争论L是从夏娃而来,其他组是登上方舟的洪水前分支。因此,我们可以从被接受的圣经叙述辩解L, M和N登上了方舟。分支超群R也可能了上了方舟,这要视舟上的女性数目而定。所有的线粒体脱氧核糖核酸单倍群都是L, 然后M和N及随后R的分支,R本身是单倍群N的突变。因此,圣经说明和现代科学脱氧核糖核酸之间的唯一争论是数学模型比圣经年表需更长的时间突变之假设。那假设是基于线粒体脱氧核糖核酸不迫使人类基因突变的前提,而那假设现已被显示是错误的。圣经的“死亡是由罪而来”声明可论证为脱氧核糖核酸系统的一个状况,如我们之前所见。从巴斯德研究院看来,线粒体脱氧核糖核酸导致突变,损坏通过疾病以及辐射发生,如我们现在所发现的。我们的脱氧核糖核酸正通过脱氧核糖核酸对人类基因的直接影响逐步突变。我们在阿当中以完美开始但随着时日转弱。我们未进化;我们退化。我们的脱氧核糖核酸暴露于不同形式的线粒体脱氧核糖核酸而突变。从逻辑而言,不同的线粒体脱氧核糖核酸必在人类基因中造成进一步的突变因而影响Y染色体。
因此,从原有的单倍群L我们得到M和N。这两组是单倍群L的独立直接突变。
我们可假设夏娃生产L系而闪、含和雅弗的三位妻子至少是三个组L, M和N。 因诺亚可能有未被提及的女儿而她们的线粒体脱氧核糖核酸系可能是L,或M或N的事实,可能已发生进一步的分支。若我们假设整个L系来自含的妻子,因L系近乎局限于撒哈拉以南地区的部落,它甚至可能是R。我们也得处理夏娃是黑皮肤的事实以及阿当的意思是那赤色的之事实。因此肤色的发展的能力是人类创造的原始特征。导致色素沉著的黑色素减少的解释是,我们离赤道更远,肤色就必须更浅才能生存。黑色素增加我们吸收的维他D会较少。因此在太阳最强烈的地方我们需要较多的黑色素以便减少吸收维他命D的能力。因此,在太阳最强烈的地方我们需要较小的吸收维他命D能力以及较多的力以及较多的黑色素保护;而在太阳较弱的地方需要更多的维他命D吸收能力和较少的保护。因此,诺亚的后代更向北移,他们的黑色素变得更浅。
闪和雅弗的妻子是L, 可能M和N或者也是R的分支。
M和 N两组形成了以下的亚组:
M产生三个分支:
M亚群包括:
C和Z, 彼此分裂,以及D和G;
E分支;和
Q分支
我们因此可推断诺亚众子的妻子是取自一个家庭世系,也在母系中保持后代的纯净。L2和L3的分裂可能来自洪水前的家庭结构。诺亚的女儿和儿子的妻子们可能有着L的所有三个分支和M, N以及可能R的基本核心亚群。
方舟上的妇女–即使只有诺亚的女儿和闪、含和雅弗的妻子–因此可轻易具有现代线粒体脱氧核糖核酸多种性的基础。即使在今天的已婚儿子家庭中发现这种多样性,我们也不会感到惊奇。在巴勒斯坦、埃及和南巴基斯坦发现这些多种性现今是寻常的。
超群M
印度人和巴基斯坦人之中发现最大的M数量和百分比。这组从那里向北和东分散至蒙古和中国汉族继而至日本和台湾。M分支成C和D发生在东方干草原汉族和蒙古人及日本人之中。
C分支在蒙古人和亚群中发生。鄂温克族主要是C,只有小百分比的M, D和U。西伯利亚中东部的布里亚特族是C和D,有一些L3然后突变G。这突变被带入东方进入尼夫赫人中,并在西伯利亚东部的伊捷尔缅人中最多产,并进入西伯利亚远东的楚科奇人之中。
M突变为单倍群E在台湾原住民中发生,而在加里曼丹的沙巴多产。Q变种在巴布亚新几内亚发生。因此,以上线粒体脱氧核糖核酸的变种在族群从印度的古实逐步东移进入蒙古、中国、东南亚和美拉尼西亚时发生。
这是洪水后M超群线粒体脱氧核糖核酸的最初演进。
超群N
超群N现在主要在中亚和东亚以及澳洲土著中发现。N组分裂为:
N亚群,单倍群I和W;
A和X分支;
Y分支;以及
R分支。
N基集中在澳洲原住民之中,与在那里所发现的人类Y染色体单倍群C有大致相同的分布。那事实显示这些人可能是从古实人向东的分布前来澳洲并保持孤立。他们的基本 脱氧核糖核酸未被其他线粒体掺入而突变。他们的基本结构未辩护长久的居留而是一个未因与其他线粒体脱氧核糖核酸单倍群通婚而承受强迫突变的孤立脱氧核糖核酸系统。澳洲的P结构可能与美拉尼西亚人的Y染色体单倍群K和RxR1一起进入。有高浓度的线粒体脱氧核糖核酸单倍群P和Q与一些M。澳洲有很小的线粒体脱氧核糖核酸 M,巴布亚新几内亚也有相同小百分比的线粒体脱氧核糖核酸N。这将显示偶尔的交换,或来自被俘虏的女人。有争论指当塔斯马尼亚原住民前来时人类Y染色体单倍群K和线粒体脱氧核糖核酸 P可能已与巴布亚人一起在那里,而且实际上是在澳洲土著的Y染色体C和线粒体脱氧核糖核酸N之前。那是不确定的,但塔斯马尼亚人属巴布人类型,与在纽西兰的毛利人之前的莫里奥里人一样。
N线粒体脱氧核糖核酸结构也在日本人、中国汉人、台湾汉人、阿尔泰人、乌兹别克人、科米人、波斯人、土耳其人中以及苏联南部发现。N超群分裂成N, I和W亚群。
I亚群相当罕见,虽然小,在库尔德人中有显著百分比的出现并或许从那里的库尔德人进入土耳其。它在埃及只有小百分比然后进入伊比利亚、法国、英国、斯堪的纳维亚和冰岛。N和I的分裂似于N组往东和I给往西之前在中东发生。W也在库尔德人之中,如U和K以及HV和H分支一样。因此,这两个分支和亚群在那里有代表,但母体单倍群类型R没有代表。N, I,和W组皆在巴勒斯坦/埃及出现。脱氧核糖核酸单倍群I可能与盎格鲁撒克逊人或较早的凯尔特迁移一起来自中东 ,可能甚至在公元前第十一世纪特洛伊败落之后。
A和X线粒体脱氧核糖核酸单倍群主要是在美印第安人之中出现。作为分支,单倍群X是在他们移入北美洲东北部时发生。
Y是在北亚发生的突变,并在尼夫赫人中发现。
超群 R
单倍群R在泰国人中发现,在印度,南巴基斯坦有最大的百分比,而在中国汉人和哈扎拉人中有较小的百分比。
R超群分成以下:
B;
F;
分裂成H和V的HV;
P;
J和T分支;以及
U,K来自它。
虽然单倍群R在以上泰国人和其他族群中发现,但 B分支较显著。B在中国汉族中发现,在中国南部有较大的百分比 ,在台湾原住民中有更大的百分比;而它也在台湾汉族和日本人之中出现。单倍群B南移入巴布亚新几内亚并在波利尼西亚中有最大的密度,超过他们线粒体脱氧核糖核酸的90%。B亚群在跨越太平洋,美国北部西岸、中部和南部发现最大的密度,显示迁移是从中亚至东南亚至波利尼西亚至美国;但美国那里的人类Y染色体不是C而是Q。阿拉斯加只有C和Q一起,然后进入美国中北部。这C 路线必也横越向北,或是人类Y染色体Q族群取了人类Y染色体C男性的女人并消灭了她们。这可能如此,因为近期一人类Y染色体C基男性的发现–被检定为北美洲西岸的一名澳洲原住民而且比任何其他发现更早-似指示了这个迁移。
A, X和D一起出现显示从北亚横跨太平洋,出自单倍群N的A分支出现在台湾和日本,并以两组向北伸展至西伯利亚远东的楚科奇人和横越阿留申人。北部族群成为了阿拉斯加爱斯基摩人,南部族群成为阿拉斯加印第安人。北部族群似乎是形成纳得内(齐帕威)线粒体脱氧核糖核酸 (主要出自单倍群A)的最初北部移民。A组朝东和北进入格陵兰,D组与A, C和B进入南美洲并进入美国东南部地区。Q男性因此有四个母系包括在北美洲以及在现是智利的地区发现的后期线粒体脱氧核糖核酸变种U。因此,美国的殖民是很晚的,其人类Y染色体和线粒体脱氧核糖核酸突变是第二和最后形成的。
HV组在中东从R突变,而R和HV皆在波斯和南巴基斯坦发现。单倍群H变种是在乌兹别克族和横跨欧洲到冰岛中央干草原的曼西、哈扎拉和科米人中发现。H形成西欧人主要的线粒体脱氧核糖核酸结构。它也在巴勒斯坦、埃及和非洲西北部至摩洛哥中被发现。单倍群V变种数量较小,除了在斯堪的纳维亚北部的萨米人之中,其百分比有戏剧性的转变。 R分支J, T, U和K的百分比在欧洲有相同的分布,除了萨米人和库尔德人之外,他们之中有显著的U,除在库尔德人之中发现的K变种以外。萨米人中没有K变种,显示他们没有变异并在它发生之前从其他U群分裂。苏联的其他北部部族凯特人和科米人有显著的U,但他们也有未在萨米人中发现的F或T变异。萨米人中也有一些百分比的D和H及一些Z出现。
附表展示线粒体脱氧核糖核酸单倍群分支的演进。
结论
在容纳,或迁就我们在此所述科学进展的情况下,线粒体脱氧核糖核酸变异和单倍群中并没有任何东西可排除圣经的故事和创世纪的叙述。
线粒体单倍群L, M, N及可能R都存在于方舟上的妇女中。各群体随着时日的推进离开中东与移动部落和家庭异种交配而产生了突变。
接下来我们将回到人类Y染色体结构以设法鉴定涉及的部落和民族,并将他们置入创世纪的叙述中。
人类Y染色体群体
所有其他人类Y染色体群体,从G到R2,都衍生自中心单倍群F。这F组对闪和雅弗的已知世系是极为重要的。我们将以F世系开始,即P14, M89, M213。这系是G, H, I, J, 和K的基系。
我们知道闪米特族的已知世系是G, I和J。也有一些E3b非洲或含米特人世系和一些R1a及R1b。这些世系和G一起也传到土耳基、乔治亚/亚美尼亚和意大利。
一般看法将中东阿拉伯人视为单倍群J,而与闪有确定清楚血统的犹太亚伦祭司是J2。这单倍群分支也确定津巴布韦伦巴部落的布巴族,他们与其余犹大/利未人分离长达二千五百年。因此这J2分支至少与这分离同样古老。
犹太教中也有相当数量的分支,显示犹太教是一个宗教而非单一的单倍群世系。
例如,亚伦祭司被确定为,阿肯纳兹犹太人的利未结构有52%是R1a1,为在苏联和中央及东部干草原以及印度雅利安人中发现的确定雅弗世系。它属斯拉夫。此外,东欧所有犹太人有25%是E3b, 显示含米特根源。
一个有关E3b根源的假设指埃及人与以色列人异种繁殖。另一个说混合众涉及E3bs,因为这群众中有大约两百万名以色列人以及大约六十万名混合众。那将构成其中的25%。不过,当我们阅读美勒休斯人的遗传说明时他们宣称也是出埃及混合众的一部分。他们是R1b。因此,有关的解释须涉及后期含米特世系皈依犹太教的发展。我们在约书亚下迦南的占据中发现这些。
迦南是含的儿子而一些部落获得倖免并加入以色列,一些通过诡计而行。埃塞俄比亚人的皈依也是重要的,因此我们足以解释E3b世系占阿什肯纳兹人25%的现象。阿摩利人和南部赫梯人也进入并与以色列人繁殖。叙利亚、土耳其也有显著的E3b水平,而在希腊人之中高达30%。希腊人有达30%的单倍群 J或已知的闪米特族以及达20%的单倍群 I,这也是已知闪米特族和欧洲人中广布的单倍群。我们将在较后讨论这些。
阿什肯纳兹线粒体脱氧核糖核酸
另一重要的方面是阿什肯纳兹犹太人的线粒体脱氧核糖核酸。阿什肯纳兹犹太人的线粒体脱氧核糖核酸被归类为近东,但我们需要看它的结构。如我们在线粒体脱氧核糖核酸方面所看到的,所有其他群组来自单倍群L3, 它分裂成两个主要的M和N组。R来自N,随着N和R的分支。N在中东分裂而形成Nu亚群即N (N1), I和W。我们知道I在库尔德人的祖先中出现,他们住在米提亚人(为玛代后代)曾经居住,现称库尔德斯坦和伊拉克及伊朗或波斯边境地区的丘陵地带。这亚群的其他组形成 N1等。
10%的阿什肯纳兹犹太人是线粒体脱氧核糖核酸N1b。这类型与祖传N1b(145-176G-223)的差别只是一个单跃迁。这可能是最古老的线粒体脱氧核糖核酸之一,除了L2之外。L2 脱氧核糖核酸如我们所看到的,是最初L线粒体脱氧核糖核酸的分支。这的根源是含米特族而现大部分是撒哈拉以南人和埃塞俄比亚人。它可能来自迦南人或混合众。摩西的第一位妻子也是埃塞俄比亚人而迦南人或也有这脱氧核糖核酸。也有M1近东类型,即M组的最初分支。其余都是R的分支,构成阿什肯纳兹线料体脱氧核糖核酸的60%。
阿什肯纳兹犹太人中的近东线粒体脱氧核糖核酸是N1b, H, J, K, L2, preHV, U7, M1, U1B (参见贝哈尔等,阿什肯纳兹犹太人口基因库线粒体脱氧核糖核酸中的差别瓶颈效应,页8,9,19; 也见阿什肯纳兹犹在人口早期历史中遗传瓶颈的线粒体脱氧核糖核酸证据,欧洲人类遗传学杂志 ,2004, 页1-10)。
线粒体脱氧核糖核酸有低百分比的,如U5a和V, 被认为来自欧洲混合(贝哈尔等,页 10)。不过,V在萨米人之中显著存在,在欧洲人中的百分比则较小,显著少过10%。
最新的分支发生在 U,分裂成K。
32%阿什肯纳基犹太人的线粒体脱氧核糖核酸是K,包含四个主要子类型。在非犹太人中最高的百分比是在德鲁兹人中,达16%。阿什肯纳兹犹太人次高的线粒体脱氧核糖核酸是H,达21%,其中7%是H祖传。布列颠阿什肯纳兹犹太线粒体脱氧核糖核酸是CRS模态单倍型。因此我们可假设HV的原有分支很可能是在这些女性(早期以色列?)的早期人口中发生。其次是J1,达7%(贝哈尔,如上)
在这方面重要的是H组成阿什肯纳兹犹太线粒体脱氧核糖核酸的21%,而那脱氧核糖核酸是整个欧洲人口主要线粒体脱氧核糖核酸序列,冰岛甫在50%之下,布列颠岛达60%,伊比利亚65%以及法国50%。它在人口东移时缓慢减少。它在土耳基人和巴勒斯坦/埃及人当中跌至25%,并在库尔德人、波斯人、南巴基斯坦人和哈扎拉人之中进一步下跌,在乌兹别克人中增加至超过22%以及在北苏联的科米人之中增达 30%。
因此可合理地假设是西欧基本的相同女性也是很多阿什肯纳兹女性的基本,以及突变在那些部族中发生。
U 至K的基本突变似在中东以色列北部和黎巴嫩的地区发生,并包括形成德鲁兹世系祖先女性的重要部分。她们是32% 阿什肯纳兹犹太人的祖先。其他有高部分K的人是库尔德人,其中U形成主要部分。U形成所有欧洲线粒体脱氧核糖核酸的重要部分。它在萨米人中最高。HV, H, U, V和K 与 I和 J是所有欧洲人的主要组。H 形成超过50%的盎格鲁凯尔特人、斯堪的纳维亚人和法国人,在伊比利亚甚至更大。阿什肯纳兹人和欧洲人之间的显著不同是,K分支在他们之中大过在欧洲,因此K的移动可能是在较后进入欧洲,或在那里从U的突变较少。很可能这些分支来自帕提亚联盟的解散和游牧民族移入在奥西之下的欧洲,它从公元第二世纪结束时开始。这两组可能形成了由凯尔特人和以色列人及来自赫梯联盟的犹太人组成的相同锡西厄游牧民族。帕提亚和锡西厄游牧民族移入欧洲而其他部落成形成可萨联盟并成为可萨帝国,在大约公元740年皈依犹太教,直到第十一世纪以及他们被拉斯所挫败并在较后与汉族一起迁移进入欧洲。这解释了为何阿什肯纳兹利未人 的非闪米特利未脱氧核糖核酸(它大部分的人类Y染色体)是R1a1,属斯拉夫而非凯尔特。
我们也应记住自塔林德时期以来的犹太身份未严格跟随圣经系统而是通过从犹太妇女的母传后裔或经拉比授权皈依而得(也见贝哈尔等,阿什肯纳兹利未人的多重起源: 近东和欧洲血统的人类Y染色体证据 , J.哈姆。Genet. 73:000-000, 2003)。这脱氧核糖核酸被认为来自可萨的皈依,以及或有达五十名男性最初登记为来自斯拉夫可萨游牧民族的利未人。似乎也有一个单一男性事件在移入欧洲之后显著扩展至s(如上)。柯希宁和利未人各由~4%的犹太人口组成。根据犹太百科全书(1972), 异教徒可成为以色列人但非利未人或柯希宁人(参见,如上)。这规则显然未在可萨人皈依中被遵守。在阿什肯纳兹利未R1a1的单倍型中,小随体单倍型紧紧聚于模态单倍型(16-12-25-10-11-13)周围,它是由单倍群内74%的阿什肯纳兹人和总体38%的阿什肯纳兹利未人组成。 这单倍型整体均匀分布(贝哈尔等,如上,也见表4)。
犹太人试图将阿什肯纳兹的根源局限于从法国东北部至德国南部的欧洲西北部小地区内。他们自称起源早至第六世纪。这地区在那时是帕提亚游牧民族解散并移入布列颠、斯堪的纳维亚、法国和西班牙及意大利的中央地点。很可能这游牧民族的一些犹太元素迁移到那里或在较后追随这民族。问题是,较早指依地语是从第十世纪发展的定义,谓阿什肯纳兹语表面上是基于希伯来字母但词语是借自德语词汇的语言。无论如何,它实际上是一个词汇替代斯拉夫方言(韦克斯勒, 1993, 参考如上)。这与索布语和贝拉鲁斯语有关。这假设现由大部分斯拉夫人的阿什肯纳兹利未人类Y染色体所证明。现在它被视为被给予希伯来字母和德语词汇的索布语。R1a1人类Y染色体也与索布和贝拉鲁斯人类Y染色体有关。这些民族可能会被鉴定为阿什肯纳兹的后裔和可萨游牧民族的一部分,因而可论证地是斯拉夫语族。
瑟法底一词最初应用在犹太人身上时,是与1492年被驱逐前的西班牙犹太人有关,在布列颠的犹太人也一样。不过,现有的用法将它应用于所有追随瑟法底崇拜仪式的北非和远东犹太人身上。
The 利未柯希宁人或祭司的人类Y染色体,与另一方面的利未人和以色列相反,是一个明确的单倍群。阿什肯纳兹和柯希宁的NRY(人类Y染色体)明确地较其他以色列人和非以色列人与对方更相关(斯哥列基等,1997;汤姆斯等,1998, 参考如上)。他们的脱氧核糖核酸模式主要来自科恩形式 (科恩形式单倍型) 单倍型频率的差异, 由六个迅速突变的小随体所确定(汤姆斯等,1998), 以及由缓慢突变独特情况多态性所确定的单一单倍群(J2)内一组密切相关的单倍型。属于这单倍型的染色体和与它有关的群集在柯希宁人中出现的频率高,但比“以色列人”中的频率低得多。这群集的模式被发现与活在二千一百年和三千九百年前之间共同祖先的后裔一致。这些发现是以不承认人类Y染色体线粒体脱氧核糖核酸曝露导致更快的人类Y染色体突变的数学模型为基础。 的确,他们也将因为柯希宁人在利未之外婚姻的限制而在这单倍型中有所限制。
科恩形式单倍型形成最可能最早的日期是公元前1900年,因此亚伯拉罕不可能拥有这单倍型。 此外,较小的日期,即最可能的,显示科恩形式单倍型是迟至公元前100年神殿时期结束时确定。它可能是朱迪亚的以东人和犹太人之间互动的结果;以东人约于公元前160年在约翰卡奴斯之下皈依犹太教。这事实或解释为何科恩形式单倍型未在拉比人口之外被广泛发现的原因,虽然我们知道绝对的事实是以色列祭司分区之中有广大的利未人口。 这二十四分区与以色列游牧民族一起离开,除了三个分区之外;他们在公元前第八世纪被亚述人所掳,并被带过阿拉克塞斯河进入成为帕提亚和锡西厄的地方。不管与哪一个游牧民族,都应有显著数目的原本利未/科希宁人类Y染色体。科恩形式单倍型在津巴布韦伦巴族群布巴族的之中的发现,显示突变成科恩形式单倍型和J2是大约发生在巴比伦被掳时期,当伦巴族群宣称他们被分开以及以色列分成十族和犹大的两族和部分利未及一些便 雅悯之后,或他们的分开是在较后和接近公元前160年,当一些祭司在安提克伊比芬斯背教之后迁至埃及并建立神殿,或随着犹大在公元约135年哈德良敕令之后以及/或巴尔柯兹巴叛变期间的分散。从族长的闪米特F发展成它的变种(例如G, I和J) 在中东和在锡西厄可能历时长达五百年或更久,并发生在公元前第一千年或公元第一世纪期间。最新的日期是在哈德良下离散的最可能日期。
因此我们可推断J 单倍群并非原有的闪米特单倍型。闪米特人因此有超过一个传自亚伯拉罕的标准单倍群。闪米特人的原有单倍群是F并至少分成I和J,及可能来自中东,帕提亚/锡西厄并进入欧洲的G, I和J 。这前提的检讨可通过参考贝哈尔等,阿什肯纳兹利未人的多重起源(如上,表A)的结果来进行。在这表中,阿什肯纳兹利未人从九个国家的六十个父系中受检测。这些国家是奥地利-匈牙利(AH) x 10, 贝拉鲁斯(Bel) x 4, 法国(Fra) x 6, 德国(Ger) x 10, 立陶宛(Lith) x 8, 荷兰(Neth) x 5, 波兰(Pol) x 7, 罗马尼亚(Rom) x 4, 和苏联(Rus) x 6。
在对六十个人类Y染色体世系作出的检测中,只有八个实际是闪米特族。其他结果如下:
已知含米特世系12
DE (xE3) x 1 (Fra)
E3b x 11 (4 AH, 1 x Bel, 1 x Lith, 2 x Rom, 2 x Rus, 1 x Ger).
假设雅弗世系40
K (x L, N, O, P) x 1 (1 x Ger)。
N (xN3) x 1 (1 x Lith) 。
Q x 1 (1 x Rus) 。
R1 x 1 (1 x Ger)。这是非常罕见的世系,过去在两名亚美尼亚人中发现。
R1a1 x 31 (4 x AH, 3 x Bel. 3 x Lith, 6 x Pol, 1 x Rom, 1 x Rus, 4 x Fra, 7 x Ger, 22 x Neth)。
R1b x 5 (2 x AH, 1 x Rus, 1 x Fra, 1 x Neth)。
闪米特世系如下,显示展示以上所辩解闪米特世系趋异的论点。
F (x G, H, I, J, K) x 1 (1 x Rom)。
I x 1 (1 x Neth)。
J (x J2) x 2 (1 x Lith, 1 x Rus)。
J2 x 4 (2 x Lith, 1 x Pol, 1 x Neth)。
这些人类Y染色体样本的影响是相当深远的。只有四个世系共有相同的J2 世系为科恩形式单倍型。只有六个共有J 单倍群。因此只有10%受到检测的阿什肯纳兹利未人被确认为有直接的犹太/以色列人类Y染色体。F和I在受检测闪米特利未人类Y染色体中占25%。
在现有理解下,令人吃惊的数目是,三分之二的阿什肯纳兹利未人类Y染色体非闪米族。将近10 %是凯尔特人,而有令人惊异的 52%是斯拉夫人类Y染色体。
这些研究也从模型单倍型的同种性总结其祖先可能只在六百六十三年前。其成长可能只来自一个人。这将与梅因次和沃尔姆斯犹太社群的设立一致(参见犹太百科全书,1972)。R1a1 人类Y染色体的星状图案显示阿什肯纳兹犹太人口在过去一千年来有迅速的成长。阿什肯纳兹利未人的多样化提示在阿什肯纳兹人与其他犹太人口分裂后出现遗传掺入(见贝哈尔等,如上,表5)。阿什肯纳兹利未的R1a1来源因此或出自讲索布语的可萨人,他们皈依犹太教并进入利未,而利未人后来迅速在欧洲扩展。
在对犹太人口进行的检测中,我们可比较瑟法底柯希宁和阿什肯纳兹柯希宁人,而我们发现甚至在柯希宁人中有含米特和雅弗世系的存在(贝哈尔等,如上,表1)。阿什肯纳兹柯希宁(AC)的单倍群J有.8684的频率,以及瑟法底柯希宁有(SC).7536,非闪米特祭司中的频率仍达14.16和24.64巴仙之间。
这些是非亚伦人及确实的非以色列柯希宁人:
含米特人
BR (前DE JR) AC占.0263,SC占.0290。
E (前E3a) AC占.0395,SC占.0435。
假定的雅弗柯希宁人
K (xL, N3, O2b, P) AC占.0263,SC占.0290。
P (xR1a) AC占.0263,SC占.0870。
R1a1,AC占.0132,SC占.0580。
在这些频率例子中,只有37%的基本犹太人口是实际闪米特人(贝哈尔等,表1)。
单倍群J是供阿什肯纳兹以色列人(AI),占.3700,而瑟法底以色列人(SI)是.3651.
含米特组是:
Y (xBR, A3b2) SI占.0159。
BR(x DE JR) AI占.1400,SI .0794。
E (xE3a),AI .2200, SI .1905。
假定雅弗世系:
P (xR1a),AI .1500, SI .2063。
R1a1, AI .0400, SI .0159。
注意基本以色列人之中的可萨皈依者比在阿什肯纳兹利未人之中的百分比要小得多。从这些统计可能取得的结论是,贵族,以及可能可萨人的卡甘和贝克的家庭被改变信仰并被赐予利未地位,以及/或这些保持忠诚的皈依者与被保加利亚人所跟随的汉族西移一起进入欧洲。异教化的斯拉夫人保持隔离或落后,被鲁斯然后被1215年的蒙古人所横扫,并于较后时在汉族和波兰人之后以索布人和贝拉鲁斯人进入。
肯定的是拉比犹太教的追随者不完全是犹太人或以色列人,而所有犹太人的63%不是含米特人就是皈依犹太教的雅弗人;除非可显示K的一些方面是闪米特而非雅弗人,它将可把 人类Y染色体序列掷入全面的修订。
K2世系中令人感兴趣的是,在亚利桑那大学资料库中的117 K2单倍群男性中,有大约 15%是犹太人。K2人类Y染色体源自中东。韦尔斯医生认为,K2人类Y染色体是古代 腓尼基人。那将意味着腓尼基人在圣经中属于雅弗,但我们知道历史上他们是与含米特人混合的以东人。因此,他们是闪米特和含米特人。与他们联盟的雅弗海上贸易民族是他施和基提,多单和雅完船只。若K2确是主要腓尼基人类Y染色体,那它显示古代海王是诺亚众子这三个民族支部贸易联盟的集成。然后它在贸易伙伴中皈依犹太教里反映本身。
闪的后代
闪的儿子是以拦和亚述;亚法撒和路德及亚兰。
我们确知以拦人成为波斯人,亚述的后代成为亚述人,亚法撒的后代成为希伯来人,而亚兰成为叙利亚人和阿拉姆语名字的来源。他们较后也北迁进入亚美尼亚。
从亚述计划的现代脱氧核糖核酸样品,现代亚述人是L, G, J和一些R1b1,但似乎检测的 R1b1在父亲方面是英国人而在母亲方面是亚述人。因此,北部亚兰(在亚美尼亚和乔治亚的地区)和亚述发展了和 G 单倍群,它在那里发现了最大的百分比。
约瑟夫在亚兰的儿子方面给了我们清楚的提示,他说在亚兰的四名儿子当中,乌斯创建了特拉可尼和大马士革,它们是在巴勒斯坦和西勒西斯利亚之间的国家,户勒建立亚美尼亚,基帖建立巴克特里亚人,以及梅萨创建现称萨勒斯帕西尼的梅萨人。从这记录,闪米特G 单倍群似来自亚兰和亚述。约瑟夫记录说亚伯拉罕首先在大马士革与一军队住在一起 (同前,惠斯顿,译,页32)。
约瑟夫提到希伯来人约坍有十三个儿子:亚摩答、沙列、哈萨玛非、耶拉、哈多兰、乌萨、德卡拉、以巴录、亚比玛利、示巴、俄斐、尤腊和约巴。他说这些人从印度的哥本河及围绕它的土地居住在亚洲。这是现代的喀布尔和它的河谷。较远朝向印度河的土地是巴克特里亚。因此,若那是事实,他们似穿过了阿拉伯半岛而且大部分继续前往印度。结果可能是约坍的复合组在阿富汗,基帖在巴克特里亚,然后路德在旁遮普。那印度的闪米特单倍群将来自在约坍希伯来人和路德的闪米特人之中发生的突变。俄斐纯金指的是约坍其中一名居住在阿富汗或印度的儿子之土地,因此俄斐地区可能是来自印度古实进入印度或甚至斯里兰卡(锡兰)。
阿富汗人和其他普什图族
关于普什图族维基百科全书说:
"根据人类学家,讲普什图语的普什图人似源自伊朗(以及多世纪来被各种其他侵入者和移民所修改)并与波斯人、库尔德人、塔吉克人和俾路支人非常相似。这些人类学家相信普什图人有东伊朗根源因普什图语被列为东伊朗方言,与其他伊朗语以外的奥塞特语有遥远的关系(进一步详情见民族语)。
讲普什图语的普什图人自称为普什图人或布什图人,视各别所讲的是南方方言或北方方言而定。这些普什图人组成主要是伊朗人、并在阿富汗南部和东部以及巴基斯坦西部发现的 普什图族核心。不过,很多讲普什图语者经与各侵入者、邻近族群和移民(如其他伊朗人一样) 混合起来,可能包括已与突厥族混合的吉勒宰人,与塔吉克人(另一种伊朗人)产生很大相互作用的杜拉尼人,以及白沙瓦北部与达尔迪奇人混合的普什图部族。在表现型 方面,讲普什图语的普什图人总的来说主要是有地中海特征的高加索人,但金头发和蓝色及青色眼睛并不少见,特别是在遍远山族之中。”
它也说:
“从公元前第一世纪至公元第五世纪,普什图人居住的地区出现欧亚人民巨大的迁移,包括亚利安人、波斯人、斯基泰人、锡西厄人、汉人,以及希腊人。后期的侵入者将包括穆斯林阿拉伯人和中亚突厥族以及蒙古人毁灭性的攻击。”
因而有R1a和R1b 及含米特C 人类Y染色体的存在。
这篇文章说,从:
"西巴基斯坦普什图人口的随机抽样,讲普什图语普什图人是与其他伊朗族群以及讲达尔迪奇语者如卡拉萨人和努里斯坦人有关的印欧人,人类学证据看起来非常可能,但此时却非确定性的。这项检测虽仍处初步阶段,仍未显示被抽样检测的一般普什图人口与在大部分希腊人、犹太人或阿拉伯人之中发现的遗传标记有任何实质关系。可能的情况是普什图人在保持他们东方伊朗人基本遗传的同时,随着时日被各侵入者稍为改变。最终需要更多研究和更广大的脱氧核糖核酸抽样检测,才能确定结果及作为讲普什图语普什图人的一般代表。 ”
这里的结论来自闪米特族全是J,如在所提及三个族群中所发现的前提。
普什图被列为伊朗人,如我们已看到的原本是单倍群I,但作为典型中亚混合人类Y染色体组合它们各不相同。
有关方面进行了一项要将第十六世纪的普什图人列为失落十族的巴尼以色列人之一以及作为约瑟的努力。这基于他们的印-朗亲缘关系和语言而被推翻。不过,坎大哈、喀布尔以及巴克特里亚地区,在印度古实的另一边,被作为波斯帝国的省份或辖地是一项历史事实,因而他们的语言受这事实所影响。他们的脱氧核糖核酸类似以拦人而且据报历史血统以及与约坍的亲缘显示闪米特族的原有脱氧核糖核酸包括约坍希伯来人非J而是F,而其原有闪米特衍生物是I和次衍生物是J, 然后H和G。以拦和约坍的所有东方闪米特人是单倍群I而不是J。因此,这些突变是在公元前第二千年期间发生而进化论者的扩大数学模型是错误的。
杜拉尼和加尔齐族被指是伊伯拉尼人或希伯来人的直接后裔,而且可能是约坍的后代。一些也坚持来自阿富汗的拉巴尼、欣瓦里、莱瓦尼和贾吉,以及来自巴基斯坦的阿夫里迪人和尤索夫寨人也是希伯来族。
路德
路德是一个不确定的来源,有两个关于路德儿子的识别和位置的解释。我们很快就会回来讨论这个谜。
希伯来人的其他位置
希伯来人的来源和位置被视为与亚伯拉罕相同,出自迦勒底的乌尔,而亚法撒的后代被视为与在底格里斯河盆地至波斯高原的亚述和以拦后代之间,在美索不达米亚平原的阿卡德族群相同。
与波斯人有关并占据以拦北部和古迪亚高原米提亚人的其他族群并非闪米特人–他们是玛代的后裔,为雅弗族并在今天被视为库尔德人。我们较后将回来讨论他们。古迪亚人或古迪人也很可能与哥德人有联系为后期锡西厄游牧民族的一部分。
赛勒斯戈登教授也坚持早期的迈诺斯人是闪米特人而他们的语言线形文字是闪族的。他对线形文字A的作品面对争议。
亚法撒的儿子是萨拉赫和希伯(希伯来名由此而来)。他的儿子法勒和约坍看到希伯来人产生分裂。约坍有十三名儿子并占据了阿拉伯半岛的整个东方地区(创世纪10:21-32)但如我们从上所见,有很多人继续进入阿富汗或巴基斯坦。 亚伯拉罕通过夏甲的儿子和基土拉的一些儿子如米甸较后加入他们。亚伯拉罕通过法勒的世系是至拉吴、西鹿、拿鹤、他拉至亚伯拉罕、拿鹤和哈兰。亚伯拉罕被召出迦勒底的乌尔并被派入迦南地。
闪米特国家的系谱将在闪米特单倍群的分布和突变方面为我们提供线索。我们很想简单地说单倍群J是闪米特单倍群而所有其他的都不是闪米特,但鉴于这些族群的分布和混合,那不可能是对的。
例如,在波斯人和以拦人的遗传结构中,突出的是单倍群I。它超越J约达10%, 并有F和G成分在内。这些群体构成波斯人人类Y染色体的一半。中东阿拉伯人也有显著数量的I和一些G。单倍群G, I和J构成乔治亚/亚美尼亚人和土耳其人的一半,而F, G, I和J半意大利人,I和J组成希腊人人类Y染色体的一半。当我们进入欧洲时,I的平衡点戏剧性增加。
G (M201), H (M69), I (M170, M258, 和P19)及J(12f2.1)似是已知闪米族世系, K(M9)为雅弗所有后代的根基。单倍群J在欧洲北部减少,但单倍群 I在西部远至盎格鲁撒克逊几乎保持高度显著然后在巴斯克人、威尔斯人、爱尔兰人和苏格兰人中减少–但仍在这些已知凯尔特人中占高达15%。
盎格鲁撒克逊人在帕提亚帝国衰落时作为部分游牧民族,从现是伊拉克地区和它以北地区的中东进入。那发生在公元第二世纪末期。他们进入西北欧地区而游牧民族在几个浪潮中分裂。盎格鲁撒克逊人和朱特人及伦巴族、丹麦人、挪威人、撒克逊人和弗里斯人相关部落告分裂但保留了类似的多样性。北法国人也来自中东。诺曼人构成一个元素并主要是R1b,但他们含有其他脱氧核糖核酸组例如I。其他是两组法朗人。其他是两组法兰克人。这些是利法法兰克人和撒利法兰克人。撒利法兰克人只可通过父系继承但利法法兰克人可通过两者继承。如我们所知道,利法是雅弗的儿子,而且我们知道这些人的贵族出身也自称是西米里国王安迪诺一世,并是特洛伊人世袭。他们被理解为是与赫克托的儿子一起留在中东族群的后代。他们以巴黎和特洛伊命名他们的主要城市。其中有很多人越过水道在布列颠定居而这些部族(例如巴黎西人)也是R1b凯尔特人。超过35%的北法国人是单倍群I和J。约有35%的盎格鲁撒克逊人是中东单倍群I,有小量的J。大约40%的英国人是中东人而看起来是闪米特人。大部分的西北凯尔特人和盎格鲁撒克逊人是R1b以及相同遗传分支的一部分,如已知的雅弗部族。明显的例外是东部,那里斯拉夫人是R1a, 而在进入西北部的游牧民族中最大的R1a分支是挪威人,达大约30%。
以色列与雅弗的联盟
布列颠的历史作品告诉我们,威尔斯人在特洛伊城陷落后从中东进入。他们在北非与也是特洛伊城陷落倖存者的康瓦尔人会合。现在我们知道它是维鲁西亚的特洛伊王国。他们的历史说他们发现玛各的儿子已在布列颠并征服了他们。他们是利法的儿子,在中东是赫梯帝国的一部分与更远东部的哈蒂和卡尔蒂结盟。
威尔斯人被称为京利人,作为以及与高卢人一起曾占据整个布列颠和部分法国。他们被局限在现是布列塔尼的阿摩利卡。他们都是凯尔特人(歌篾人歌蔑)。他们是再侵犯安纳托利亚并建立加拉太的西米里族的根源(即歌蔑人)。他们形成凯撒呈献给希律王的四百名克娄巴特拉女王护卫。他们可能是土耳其R1b的来源,达>15% ,在叙利亚较小。土耳其人也是乌拉尔阿尔泰语族群的一部分,因此一些可能出自那来源是R1a和R1b。这两组和 Q非常接近,而这些族群之间的分支因非常小的突变而中断。因此,印第安人是凯尔特人和斯拉夫人的远亲。
爱尔兰
爱尔兰早期的居民是涅梅德人,然后是弗摩里人、袋人(或皮革人)以及丹努之子。
早期的后裔来自西班牙并被列为加德利亚人,戈达尔的后代,从雅弗起第六,戈达尔的后代(同前)。加德利亚人是来自从波里昔尼斯扩展至马萨革泰人(更大的哥德游牧民族)以及从斯基泰人(因而锡西厄人)至里海的锡西厄人。他们是游牧民族(同前,页 61)。
从遗传上我们可以确定他们为歌蔑和玛各的的儿子。人类Y染色体的变异显示阿什肯纳兹犹太人是较R1a1多于R1b,因此这些变异必是在这些族群分支后发生。所有阿什肯纳兹利未人的52%是歌蔑的后代。因此,关于歌蔑的预言与今天的以色列有直接关联。
非洲,特别是喀麦隆发现一个古老的RxR1人类Y染色体种类。它可能源自雅弗族进入非洲的部族迁移,或在澳洲原住民RxR1族群跟随他们之前的古实单倍群C游牧民族前往澳洲时从东方进入,或甚至迟至从西班牙入侵北非并于较后被击败的汪达尔人。
我们从爱尔兰的历史和世谱中知道米勒休斯人是玛各的后代,他们的检测是R1b。不过,我们知道他们在相当迟的时期进入爱尔兰及取代先前在那里的一个部族。那部族统治爱尔兰数百年并以达南神族著称,称为丹族或达南人。据说他们也与非利士人、早期希腊人和看来腓尼基人一起航行(达南人和但族–希伯来人是希腊人吗? BAR, 1976年六月)。很有可能这个部族是爱尔兰出现单倍群I的原因,因为盎格鲁撒克逊人未在他们进入布列颠时占领爱尔兰,而诺曼人主要是R1b及一些来自盎格鲁撒克逊人的I,但I的百分比比例不代表在爱尔兰的百分比。I的比例几乎与在苏格兰的I比例同样高,但那里的主要人类Y染色体也是R1b。这两个地区可能有来自挪威I的注入。若历史记录和它的解释说达南神族确是以色列但的旁系子孙,那我们有一个明显的链接可显示北部部族是单倍群I。因此40%的盎格鲁撒克孙人将是闪米特人而且可能是以色列的,如记录所断言。针对这方面的争论是,少过2%的犹太人是单倍群I。然而,当我们考虑到只有37%的犹太人是闪米特族,那将样品提高至已知犹太人的大约6%,因此很可能代表留下或返回犹大其他部族的残余。
也有一个来自他施船只的已知腓尼基K2出现在推罗、马尔他及威尔斯。美国总统托马斯杰弗逊自称是威尔斯后裔并属这K2 单倍群威尔斯子分化体。
爱尔兰的第一个殖民地是由巴托兰所管理,他属于希腊人的古代世系,在那之前,希腊 土地被侵略并由来自非洲的E3b含后代所居住。第二个殖民地是由涅梅德掌管,他战绩良好巩固了他的殖民地。不过,弗摩里人侵入并在科克击败涅梅德人而接管了爱尔兰(麦克盖根和米歇尔, 爱尔兰的历史, 1867,页54)。
袋人被麦克盖根鉴定为比利时人,而在布列颠有一个比利其族。弗摩里人或被击败,或离弃这土地,袋人占有了它。他们被达南神族所取代,后者是涅梅德人的后裔,他们通过德国和取道挪威及丹麦返回爱尔兰(麦克盖根, 如上,页55)。
麦克盖根宣称他们可能是康沃尔顿诺尼亚的后代。他说他们是涅梅德孙子约巴的后裔,约巴率领源自爱尔兰的部分殖民进入北德(如上, 页55)。因此,爱尔兰的原有殖民是古希腊与涅梅德后代,即达南神族或但族的联盟。这些是早期的航海但族与古代海王的结盟。
麦克盖根也宣称涅梅德通过菲格斯所生的孙子贝雷登莫尔在布列颠确立本身和他的部族,根据卡舍尔诗篇以他的名字见称–据报导他们居住在那里为布立吞人。麦克盖根说那至少与蒙茅斯杰弗里的说明一样可信。这将说明苏格兰R1b 皮克特人之中I单倍群(岛屿亚分支)的分布。他们未来自挪威并几乎是布列颠岛屿所独有的。皮克特人也是布列颠早期的殖民,事实上比苏格兰人早一千年。
同时,或是因为弗摩里人被击败或离开,另一批五千入进入并将爱尔兰分割成五大牧首区,以伦斯特的总督史连基作为史连基、罗立(路瑞)、甘、甘南和仙敢五大牧首区的首长–涅梅德族的代拉的所有孩子。这些人当时以三个不同的名字见称:加里宁人、顿诺尼亚人和比利时人。他们在九个国王之下统治八十年,最后一位国王是伊奥卡。他娶了一名西班牙王子的女儿。她被埋在以她为名,米斯的代尔登。
比利其人也位于最接近英国顿诺尼亚的地点,这三个名字很可能提示这三个从欧洲移至英国至爱尔兰部族的联盟。加里宁人似是高卢人,因而也是迦拉太人。无论如何,很可能通过达南神族下的涅梅德人我们看到单倍群I与凯尔特的R1b(AMH)一起侵入。来自丹麦的这一族群比后期帕提亚游牧民族的R1b 丹麦人来得更早(后期的丹麦人是DYS 390/391 23/11, 而丹麦共有两个不同的I组,一是盎格鲁撒克逊类型;见下)。这项迁移也可能与汉族或甚至在他们之前来自公元前第二千年的船运族群一起进行。那时波罗的海有广泛的琥珀和贸易系统。如在以上所看到的,我们知道K2他施/腓尼基商人在威尔斯和康沃尔,而且这仍在那儿存在。
达南神族是从这时起在努亚卡艾里奥兰之下抵达爱尔兰,他打败了在马约郡伊奥卡之下的袋人。这些袋人被赶出去并在北部岛屿寻求庇护(如上, 页55)。这些定居在这些岛屿的人是 R1b。此外,有I1b2a–岛屿亚分支的存在而这单倍群几乎仅在布列颠群岛以及大量在苏格兰发现。它不是挪威的维京人。相同的I单倍群在巴斯克人中发现。看来它肯定未来自斯堪的纳维亚而是从中东进入西班牙而其中一些留在巴斯克人之中的达南神族。主要部分继往爱尔兰并从那里前往苏格兰和群岛。他们也在皮克特人中被发现,因此他们必已在苏格兰人从爱尔兰迁往阿尔巴之前已到了那里,因为苏格兰在九人质的尼尔重新命名它之前已被称呼。他们未与苏格兰人或米勒休斯人一起从欧洲进来。但族作为达南神族的意义是它并不是很多布列颠以色列人所认为的局限于爱尔兰。这族群集中在苏格兰也有较小数目在威尔斯人之间,一些在英国以及或者在巴斯克人之中。从启示录章7中我们看到但和以法莲停止存在为单独实体而合成的约瑟再合并,但这次没有隔开的玛拿西。
这些岛屿是苏格兰海岸的岛,因为图勒或冰岛的任何殖民地化是在数世纪后才发生。
米勒休斯人是取道西班牙前来爱尔兰,而一亚族从西班牙进入英国并以布里甘特斯人见称。他们在目前大致是兰开夏地区的西英格兰定居。
爱尔兰人–直到米勒休斯人约于公元前500年进入时,如我们在上面所见–是由三个称为弗摩里人、袋人和达南神族或但族的族群组成。
我们确知米勒休斯人是R1b凯尔特人,而我们有他们的系谱和他们的脱氧核糖核酸来与这些血统匹配。这些凯尔特人符合其他凯尔特R1b人类Y染色体序列但因地区和部族而异。
西欧R1b凯尔特单倍群的主要决定因素在R1b1单倍群分支中发现。R1b的四个亚分支是衍自DYS标记390和391。
390/391有24/11数值的大西洋形式单倍型(AMH)在从西班牙至苏格兰的大西洋海岸发现。这些凯尔特人是欧洲那些较年长的居民并通过较早的分支传自雅弗,他们在巴斯克人中被发现。
盎格鲁撒克逊人从公元410年罗马占领布列颠之后以及在诺曼征服之前进来。他们有R1b和I单倍群。盎格鲁撒克逊的R1b(R1b1)亚分支由具有23/11数值的DYS390和391标记来认别。这观点开放讨论而且可能不是一个显明的鉴定。这些数值在西北欧游牧民族在弗兰德(比利时)、荷兰、西北德和丹麦确立的其他民族中发现。弗里斯人和盎格鲁撒克逊人等皆共有这识别标记。他们和德国撒克逊人实际上是相同民族。
The other 亚分支subclades of R1b的其他亚分支是24/10的重复,在欧洲各地发现,以及 23/10的重复,在东南德发现并进入奥地利。因此他们都是来自凯尔特人或原本来自锡西厄和帕提亚歌蔑后代的不同世系。
在拥有I1a盎格鲁撒克逊人之中所发现的单倍群I变异是I1aAS(1-8)。I1aAS是I1单倍群中最常见的变异,并以它在盎格鲁撒克逊人从中东迁入后原本居留的地方为名。它在我们进入东南德及更远的地方中散去。可能这单倍群的变异标志着十族中一些部族的祖先测定。这些民族不变地与上述赫梯凯尔特人互相联系。 AMH是一个较早的组,与在R1b1和I1aAS两方面都有明显标记的盎格鲁撒克逊人不同。
对丹麦人Isles I1aAS的测定与在这大陸的德国祖先记述对比将产生更多的资料。I1a-(诺斯)是在瑞士和丹麦发现的最稠密类型,挪威紧随其后。在欧洲大陸波罗的海和北海只发现小数量,次于丹麦的I1a–AS。因此,丹麦的I 组混合,但丹麦人主要是盎格鲁撒克逊类型(见文章Y-单倍群I之中和它们扩大模型单倍型(以及R1b种类)的人口变化,肯诺特维- owww.northwestanalysis.net)。
I1a uN (超诺斯)在挪威达到它的顶峰。它因为DYS 385 b 从14到15的一个变换而与I1a有异。它也基于DYS461的重复数值而分裂成两个类型。I1a uN1在 DYS 461和449有12, 28形式,在大部分I1a方面是普遍的。I1a uN2有11, 29在DYS 461和449中重复,与挪威的联系较瑞士和丹麦强。这些分支可能是在公元第三和第四世纪斯堪的纳维亚的大迁徒时期之后发生。
I1a1 (old I1a4) 是I1a的小亚分支,在东欧的 I1a发现10%。
I1b1 Din (狄那里)之名取自巴尔干的一座山脉接近它最多产的地方。 它也在东欧的很多地区散布。
I1b1–西(西方)是(旧)I1b, P37.2 + 的变异,在德国的波罗的海和北海海岸地区大量发现然后传至布列颠岛屿。它是由DYS 388的十五个重复而平常非狄那里I1b1的十三重复显著鉴别。这类型最近也在爱沙尼亚发现。I1b1-Isles是近期发现的I1b1变异,它似主要留在布列颠岛屿上。鉴定标记是 DYS 385, 463, 和464。
I1b1a (旧) I1b2, I1b1 M26+的亚分支在萨丁尼亚人口中有很大的百分比并传入伊比利亚。它在欧洲北部未有任何显著的发现。
I1b2a–Cont(大陸) (旧I1c)是I1b2a的主要变异。它在西北德、荷兰至进入丹麦最稠密,一些在瑞典南部和挪威发现。
显著的数量也在布列颠群岛的日耳曼和斯堪的纳维亚的移民/入侵者中发现。
I1b2a-Root是I1b2a一个不寻常的变异,从意大利和伊比利亚传至丹麦并基于四个标记分裂成两个类型。
I1b2*是从意大利和伊比利亚至丹麦发现的另一个类型,主要基于DYS 448标记分裂成两个类型。
I1b* (旧I1(y)是这树中的一个小类型并有10/10 DYS 459a,b为它的形式特征,在YCA11a,b有19.19或19.21以及在DYS389i,ii 有12,29。
这些I 单倍群亚分支类型被认为可鉴定希伯来人而R1a 和R1b分支将鉴定赫梯联盟的分枝;这联盟是在锡西厄和帕提亚及干草原中成立并渐进欧洲。
单倍群I和J现被争论并被接受为F的亚群,它曾是并合IJ组,为一个民族。
从中东及锡西厄殖民化欧洲
以色列约于公元前720年在亚述人之下被掳,并被带入北部进入后来成为帕提亚和锡西厄的地区。他们在那里与形成北部赫梯族的凯尔特人建立了联盟。
赫梯和锡西厄游牧民族的一些成员在特洛伊城陷落后进入欧洲和布列颠及爱尔兰和欧洲其他地区。
在奥西之下,以及在他们于现纪元的第二世纪中在帕提亚被波斯人击败后,帕提亚和部分锡西厄游牧民族迁至西北欧并建立了一些民族,包括盎格鲁撒克逊人、朱特人、伦巴族、弗里斯人和荷兰人,德国撒克逊人、丹麦人、挪威人和一些瑞典人、伦巴族、哥特人、苏维汇人、达尔马提亚人、汪达尔人和他族。他们建立了西欧国家,从西班牙和葡萄牙至丹麦及挪威,横越至布列颠和图勒(冰岛)和格林兰。
斯拉夫人最后迁入,因此有关的分去是西欧R1b之下雅弗的后代和东欧R1a之下雅弗的后代,他们都与希伯来闪米特族和亚述的后裔混合。
罗什、米设和土巴的斯拉夫人全是R1a, R1a1, 斯拉夫和斯堪的纳维亚人。苏联王国是由斯堪的纳维亚的瓦兰吉人鲁斯所建立,他以公元850-950年的基辅和诺夫哥罗德为基础建立王国。
从锡西厄到爱尔兰
涅梅德是达南神族的祖先。占领爱尔兰的定时是有趣的。
麦克盖根报导古代历史说,锡西厄国王芬尼斯法尔萨有两个儿子,尼诺尔是长子和王位继承人。
次子是纽尔,他精通因巴别塔的混乱而成倍增加的语言。纽尔航往埃及,在那儿与埃及法老(辛克里斯)的女儿结婚 并在红海连界的卡巴西仁土地上确立。纽尔通过苏格达生下一个儿子戈达尔。据说他在出埃及时期被一毒蛇咬着并将它呈献给摩西。那被咬伤口转青而这标志使他取得戈达尔格拉斯或戈达拉斯之名(格拉斯在以他母亲世界苏格达为名的苏格兰语中的意思是青)。
他们因此是出埃及中混合众的一部份。米勒休斯族谱似也作此索取。
他们定居在法老分配给他女儿后代的地区。经过三代后他们被法老恩杜尔下的埃及人攻击而被迫迁移,在他们的领袖戈达尔之子伊苏之子苏尔的领导下前往革哩底;苏尔在那儿去世,由他的儿子伊伯尔继位,他也以希伯-苏格见称(麦克盖根, 如上, 页56-57)。
希伯-苏格从那里迁移,取道爱瑟海和静海前往锡西厄的塔奈斯河。他们在那里在艾纳蒙、泰特、艾德农和兰菲恩之下的锡西厄人当中住了四代。
锡西厄人远远传至在公元前第二千年是现有中国的维吾尔族人自治区。我们发现了他们 保存良好的木乃伊和格子花呢短裙,很多类似苏格兰人的狩猪格子呢。
加德利亚人被怀疑和被迫害,并在艾德农之下的阿马逊人中寻求庇护,然后在他的儿子兰菲恩之下离开前往格杜雷。一些人认为这是哥瑟亚,但根据奥弗莱厄蒂它是在非洲的格杜利亚;他们在那里在八个国王之下逗留了八代,这些国王是兰菲恩的后代;最后一位是比尔,为加兰赫或称为米勒休斯(Mileag-Espaine-拉丁语)的父亲,米勒休斯是米勒休斯人的祖先或古爱尔兰人。现在我们知道阿马逊人是锡西厄族的已婚妇女,她们与他们的男人一起骑马战斗。
米勒休斯返回锡西厄短时期,他受到通过长子南努尔系出芬纽斯法赛的国王利弗罗的迎接。他委任他为将军并将他的女儿锡奥嫁给他,给他生了两个儿子多恩和艾利兹,姓弗阿布洛。在他的妻子死后,他与他的岳父产生不和,与他的儿子和他的加德利亚人军队前往埃及,并在那里受到部队指挥参与与埃塞俄比亚人的战争。
他被赐予法老涅多尼巴斯的一位妻子,也名叫苏格达,为他生下两个儿子赫伯-菲昂和安河金。
在那时期里,他安排了十二位年轻人接受艺术和科学教育以返回西班牙。
他在埃及逗留七年后返回西班牙。加德利亚人在几年大旱后离开前往爱尔兰。
因此我们看到锡西厄人和爱尔兰及苏格兰人之间的的链接和关系。P和Q布立吞人的语言是不同的,但他们与圣经人物歌蔑和玛各有历史关系。
这时帧也使我们能够确定另一个看来是涅梅德人或达南神族的人类Y染色体链接。米勒休斯爱尔兰人有R1b基本人类Y染色体结构,这在凯尔特人方面是普见的。它也有I人类Y染色体单倍群并看起来是源自闪米特族。
这相同的单倍群也在盎格鲁撒克逊人和欧洲人之中出现,一般上高达40%的水平,如我们也在克罗地亚人和挪威人中所见一样。然而,这两个民族有主要的斯拉夫单倍群R1a而非凯尔特R1b,虽然它在两个族群之中存在。
通过研究历史和脱氧核糖核酸结构我们能够将部族的移动拼凑在一起并建立链接,否则那是不可能的。
这闪米特单倍群I如何进入欧洲人中呢?
答案可在圣经中找到。
基土拉的儿子在出埃及之前迁徒北部,他们也成为斯巴达希腊人,如我们从约瑟夫得知。这些人是那些继续前往爱尔兰者,而这族群的一部分形成了涅梅德人或达南神族群体。达南神族被认为与希腊人相关,而且但人可能与基土拉的儿子们有关系因此他们解释了希腊人中的I单倍群以及一些J单倍群。E3b是较后来自非洲腓尼基迦南人的侵入。
这些人也从中东涌入。克罗地亚人有很高水平的I单倍群,百分比与斯堪的纳维亚一样高。因此在巴尔干半岛各地我们看到有很高的I单倍群人口来自不同根源,但在祖先方面有很多共同点。
赫梯联盟
大卫王从他事业的早期起就与推罗和西顿的北部王国以及赫梯族结盟,两者都是利法的后代和玛各的后代。他们因失去首都在特洛伊的维鲁西亚王国而变弱。因此,阿什肯纳兹的后裔这时曝露于以色列而以色列人加入他们并在较后使他们皈依。
阿什肯纳兹的后代和利法的后代,以及陀迦玛的后代都是歌蔑的后裔。然而玛各也有R1b。因此,R1a和R1b的分支是在这些部族分裂之后发生。一些成为R1a而另一些R1b。阿什肯纳兹的后代成为R1a, 否则R1a组称本身为阿什肯纳兹人即使他们是斯拉夫人,从他们的可萨接触看来那是非常可能的。
扫罗巩固以色列地区,但大卫将它扩展至幼发拉底河。
于公元前第十一世纪开始时,他是在加帕多加和那里的北部进入锡西厄在米设和土巴之中。我们从诗篇知晓这点(诗篇120:5)。
圣经中有两个米设。一是雅弗的儿子;另一个是闪的儿子亚兰之子,住在叙利亚。很可能大卫是在叙利亚亚兰的儿子米设众子之中,他们被视为与基达的儿子一样。
米设和土巴综合鉴别雅弗的而非亚兰之子。
在一千年前被米设接管的土耳其加帕多加地区,原称为库斯,以含的儿子和宁录的父亲古实为名。赫梯帝国国王的首都在那里,巴比伦人称他为库斯之王。
赫梯帝国被划分成北部和南部。他们是由含的后代和雅弗的后代组成,两者皆属歌蔑和玛各及玛代或米提亚人,而他们的贸易联盟是基于及与西顿一起,来自他施以及和黑斯的后裔。荷马在伊利亚特史诗中说,特洛伊群众会讲很多种语言。
推罗和黎巴嫩的居民成为雅弗之子(雅完他施和以利沙之子,以及基提和多单或罗单)海事联盟的后裔K2人类Y染色体。这些不属于黑斯的后代,我们今天看到他们是在黎巴嫩和马耳他的后裔之中。
单倍群H
单倍群H大部分局限在印度北部和南部,以及于较小的程度上在乌兹别克族和柯尔克孜族北部。南部较大的出现显示它于亚利安人侵入之前已在那里。若这是闪米族单倍群我们得鉴定突变进入这在印度孤立单倍群的闪米特民族。
合格的闪米特族是路德儿子的部族。他们不应与是麦西或埃及后裔的路低人混淆一起。 约瑟夫视这些人是占领在安那托力亚或现代土耳其的爱奥尼亚地区之吕底亚人。这说得通,因为当时他们是在亚兰西部和亚述及亚法撒南部闪米特族之中以及含族的北部。这部
分有更多资料,但他们和他们的后裔不是单倍群H。
现是伊拉克的东北部地区有个称为卢卢比的部族,他们向阿卡德人开战。他们被认为是现被库尔德人称为卢卢的氏族,但那不是相同的人,因为库尔德群体是雅弗的玛代之子或米提亚人-但他们可能已经随着时日被吸收过去。
他者鉴定路德的后代移向亚兰东部。那可能将他们置于印度的地区。房遮普里的古城是卢迪安纳,似以路德命名。有关的解释因此是,大部份族群分裂并东移而建立印度的城市。
它也解释为何印度河盆地的哈拉帕和摩亨佐达罗古城在公元前大约1750年与来述人有接触并采纳了他们的崇拜系统。我们预期在那里的古实后代中发现的C单倍群被南部的其他 H, L和J组所超越,南部族群在北印度有较大的亚利安人元素R1a, 德拉维人较古老的RxR1比南部的R1a来得多。古实的后裔向北和东迁移,印度因此是所有三个世系的结合。
亚伯拉罕的后裔
亚伯拉罕有一些后代以及部族来自他的侄甥洛特,与他有关系。洛特建立了现是约旦的摩押和亚们。以撒有两个儿子雅各和以扫,以扫的儿子建立了以东并成为与腓尼基人成为贸易系统伙伴的以东人或以土实人。他们后来在公元前160之后的马加比纪元中被约翰卡奴斯改变信仰犹太教。他们被吸纳入朱迪亚成为南部犹太人与加利利人相反。
雅各有十二族:由拉结所生的约瑟和便雅悯;拉结的侍女辟拉所生的但和拿弗他利;利亚所生的流便、西缅、利未、犹大和以萨迦;利亚的侍女悉帕生了迦得和亚设。我们知道在利亚所生的儿子中,利未发展为J2系(超过一千七百年)。
不过,达南神族的爱尔兰元素要成为但族,他们得是单倍群I 。(另一个解释可能是他们已完全根绝而单倍群I元素是来自挪威和英国。对这观点的反对是,R1a的百分比得与在苏格兰的一样大,但它相比之下极小。因此,那里的单倍群非来自斯堪的纳维亚或英国,而且也不是米勒休斯因此是达南神族。) 以色列的其他后裔很多也将有单倍群I,因此突变不但将在妻子之中发生,也将在相同母亲的儿子中发生。 另一个解释是这些突变发生在埃及而这些族群在那里分岔。约坍希伯来人和以拦人及基土拉的子孙之中必也发生这些分岔。
从以拦人和阿拉伯人之间的脱氧核糖核酸比率,看来较早的脱氧核糖核酸在这些后代中独立发展,从F进入I,为主要的单倍群,并也进入J 和G。
据记录[参考以下] ,基土拉的儿子是心兰、约珊、米但、米甸、伊施巴和书亚。约珊的儿子是示巴和底但。底但的儿子是亚书利、利都是和利乌米。米甸的儿子是以法和以弗、哈诺、亚比大和以勒大。
我们知道米甸人面对战争,首先是于以色列在埃及被掳之前后来在出埃及之后归因于偶像崇拜。他们与以实玛的儿子和在他们之前的约坍希伯来人一起占据阿拉伯半岛地区。
从约瑟夫的威信,我们知道斯巴达人是基土拉的后代。因此希腊的大量人类Y染色体单倍群I可能来自基土拉的后代,而J后来从在小亚细亚的征服以及较后世纪中的阿拉伯人进入。地中海一带有显著的闪米特单倍群J人口,但I在欧洲各地有较大的出现。希腊也有在非洲以外最大百分比的E3b人口。
若单倍群如所看来的确是闪米族,那保加利亚人和罗马尼亚人是欧洲拥有最大百分比的闪米特单倍群者。然后,西北欧和斯堪的纳维亚并未远远落在他们后头。盎格鲁撒克逊人从中东前来时带来一个基于闪米特族的历法系统,称为阿尔马纳中“计数历”–而我们的单词年鉴 是来自那个系统。剑桥保存着以月朔为根据的原有五十九天隔月年鉴。
看来帕提亚游牧民族是由凯尔特和闪米特游族民族合成。他们成为较小部族的人,迁移并重新结合住在欧洲地区。他们的语言是条顿语而且他们吸纳了在他们之前的凯尔特人,但从他们在中东成立的赫梯联盟,他们本身有很大的凯尔特比例。
现在让我们看看似来自K单倍群的分支,雅弗部族似传自它。
雅弗的后裔
单倍群K, L, M, N, O, P, Q和R看来是雅弗部族。神说他将祝福雅弗。他被告知神将扩大他以及他将住在闪的帐幕里。因此我们预期在相同的族群中找到雅弗和闪的混合。因此单倍群I和R1b及R1a组合都符合这些准绳。
K是从L至R2所有附随单倍群分支的根本分支。
大部的K是在巴布亚新几内亚和美拉尼西亚及中国汉族中。维吾尔族、乌兹别克人和蒙古人以及波利尼西亚人之中有较小的百分比。一些K也在意大利人之中发现。因此,那根本分支向东渐进,一小数目留在地中海希腊和意大利中。其他群体从那单倍突变而来。
这些是雅弗之子:歌蔑和玛各、玛代、雅完、土巴、米设和提拉。
歌蔑之子是阿什肯纳斯、利法和陀迦玛。这些是亚述人所提及的歌蔑人;亚述人是希罗多德的基米里人。他们是凯尔特人的祖先。阿什肯纳兹人形成了可萨游牧民族。利法人形成维鲁西亚的西方或特洛伊凯尔特人。他们从公元前1000年起成为古布立吞人,取代或征服在布列颠的玛各凯尔特人。盎格鲁撒克逊人和朱特人从现有纪元的第五世纪起反过来取代了他们。他们在第十世纪击退了汉族对布列颠的侵略,汉族的亨伯王在那里以他为名亨伯河的战役中被杀死。
玛各在以西结书38:2,6中与歌蔑有关。玛各因此也是凯尔特人的祖先,而且他们是后来在公元前第四/第五世纪侵略爱尔兰的米勒休斯人和在公元第五世纪进入苏格兰的锡西厄凯尔特人。约瑟夫说,玛各是锡西厄人的祖先(J的A,卷1, 章vi)。 这些群体的凯尔特语的区别称为P和Q布立吞语;它们不是相同的语言。不过,它们来自安那托力亚和西部过去是锡西厄的地区。赫梯联盟有很多语言,希腊人形容特洛伊群众的建立为有很多语言。
我们也从英国的历史记录中知道巴克斯人是现为西班牙北部伊比利亚地区的原有居民并被指是哥德尔的后裔。哥德尔或哥德尔人传自雅弗。这些历史也说他们是雅弗的儿子和孙子,土巴和他施的的后裔(天主教百科全书,卷XIV, 页176)。他施是雅完而非土巴的儿子。因此这些历史坚持西班牙有两个据称是雅弗的族群。圣经确定他施(历代志下9:21; 20:36,37; 诗篇48:7; 以赛亚书2:16; 23:1,10,14; 60:9; 耶利米书10:9; 以西结书27:12,25; 38:13; 约拿书1:3; 4:2)。这些是伊比利亚人,被划分为北部的巴斯克人和南部的塔提苏人(如上)。凯尔特人将他们本身附加在这两个较早的或原住民人口之上。然后凯尔特人在北部和西部,伊比利亚人在东部和南部,以及中部阿拉贡和卡斯提尔的混合“凯尔特伊比利亚人”。无论如何,他们之中也有I元素的存在,虽然比在爱尔兰所发现的较小,达大约>7%。腓尼基人也在那里的加的斯海岸,以及卡特亚、不尔佩、马拉加和塞克西建立了殖民地;加的斯是他们的首要城市。罗得岛人也于公元前大约910年在克雷乌斯角南部的罗萨斯建立了一个殖民地,以及佛西斯人在安波宁。迦太基人于公元前第七世纪在巴利阿里群岛定居。他们在第六世纪协助腓尼基人对抗塔提苏人然后接管主要城市加的斯并始在贝蒂卡进行贸易。他们在第一个布匿战争西西里面对损失后开始攻克西班牙,但在第二个布匿战争过后完全失去它。
巴斯克语被指属于乌拉尔-阿尔泰语组,因此与芬兰人和亚洲阿尔泰语、蒙古、满族-通古斯人和日本人有关。然而巴斯克人是R1b,一些是I。
以上关于伊比利亚的评论显示土巴建立了伊比利亚人,而约瑟夫说(如上)他们是托贝尔的后代– 因此土巴和托贝尔被认为是相同的。米设的后代原在加帕多加(如上)。后期的伊比利亚人是西哥特人,他们于公元第二世纪与游牧民族一起进来。
我们已确定米提亚人为现代的库尔德人,布林格坚持这是事实。他说,他们是亚述人纪念碑的阿马达或阿马代(约公元前840年),因此现在是在亚述东部的库尔德族。
陀迦玛被形容为是在北部最北端的部分并看来是单倍群N和它的变异。若这是事实,陀迦玛的后代将是随着N分裂成N(LLY22)后来自K的M214分支。华人和马来人是(M175)。乌拉尔-阿尔泰组和中国汉族因此是雅弗的后代,有很大的单倍群单倍群C 和D古实蒙古人与她们女性的混合。
雅完的儿子是:以利沙、他族、基提和多单。圣经说这些是异教徒岛屿在他们土地的划分;各随各的方言,宗族立国(创世记20:2-5)。
这些族群就是与推罗、西顿和迦太基的E3b腓尼基人形成古代海王的贸易帝国者。克里特岛人的祭司的肤色比其他的浅,如我们现在从克诺索斯皇宫的接班兵中所看到,从革哩底博物馆的耳环看来,他们也崇拜金牛犊。地中海的海岸周围有显著E3b的百分比存在。
雅完的后代被确定为楔形文字和阿玛纳山碑的爱奥尼亚人。希腊人最初以这名字见称,但希腊人现代的脱氧核糖核酸分支显示我们是在看着一个与原有爱奥尼亚希腊人不同的人。
以利沙的后代被理解为是埃及人的阿拉西拉–换言之,是奇里乞亚海岸的居民。布林格认为,他施可能是大数。不过,他施是在伊比利亚或现是西班牙的南部。在古时,跨大西洋商业大部分是在这港口进行。基提是从塞浦路斯运作古代海事贸易帝国的祖先。多单被理解为是罗单(历代志上1:7, 参见七十贤士译本)。
米设和土巴相互关联。塔巴里与土巴被联系一起。米设与亚述纪念碑的穆什卡有关联。我们明白这些是苏联人,而他们的单倍群主要是R1a,在苏联北部有小数量的R1b和显著的N, I和J, 以及一些E3b。
这些族群原本是在小亚细亚北部玛各人之上,但随着赫梯人被击败以及特洛伊城陷落后,他们进入了加巴帕多加的安那托力亚。这是大卫王与米设(以及或土巴)在一起时所提及的冲突。他于公元前大约1000年书写时这些部族是在南部更远的乔治亚/亚美尼亚。乔治亚一带的地区也从伊比称为伊比利亚。这可能是这名字的来源,并解释这名字在西班牙的使用。
大卫提及那一点时无疑是在设法遏制米设(或可能土巴)因他要和平他们要战争(诗篇120:7)。他也提及基达的帐幕。以西结书38:2叙及罗什、米设和土巴的领袖。罗什的意思也是主要或卓越的,因此它被译为主要领袖或王子而非一个名字;但它可能是鲁斯名词的来源。维京鲁斯从公元850-950年在基辅和诺夫哥罗德一带成立了鲁斯的瓦兰吉王国。瑞士和挪威比西欧任何其他地方有更多的R1a元素,但斯拉夫人是真正的R1a族群,波兰人有最大的百分比。罗什与苏联及穆什卡市(莫斯科)有联系而托搏尔斯克则与这些部族有联系。他们在公元前第一千年的开始被遏制之后迁往北部。然而,他们也有显著的I 单倍群人口。当以西结提及歌革以及罗什的王子米设和土巴时,他说他们是在玛各的土地上。在古代,那地区是在安那托利亚北部和特洛伊地区东部,现是土耳其、亚美尼亚和乔治亚。
提拉是特拉斯(来自提拉西亚)的原有居民,但传自他的部族似乎未留在那儿因为它的单倍群看起来是闪米特族。
有一个单倍群从单倍群P(M45, M74, P27, 92R7, P69?)发展:即Q (M242, MEH2, P36)。那是在单倍群R超群之后。Q在苏联的库普人和阿尔泰中出现,然后直接进入西伯利亚及横越太平洋进入美国。还有一个这族群可能迁移的方法,即横越大西洋。
我们现在知道的事实是,埃及和美国之间的跨大西洋船运约于公元前1000年定期进行。近期对柏林博物院木乃伊的法医检验显示这博物院中的每具木乃伊都有可卡因和烟草的消耗。英国博物院怀疑这些结果受到污染而对他们的所有木乃伊进行试验以作比较,他们未在他们的木乃伊中找到古卡因,但每一具都有正面的烟草消耗。 这些产品只在美国种植。这些木乃伊都是大约在公元前1000年时期。这贸易似乎从公元前第二千年持续至公元第一千年,如赛勒斯戈登从这证据所显示。这贸易是横越太平洋和大西洋。
美印第安人或如上所述般迁移,但他们在大卫王至罗马人时期肯定横越大西洋与伊比利亚他施人和古代海王进行贸易。有证据显示罗马于现纪元的第一世纪在那里进行贸易。据指巴尔科赫巴的起义者也在现纪元的第二世纪前到那儿,他们的硬币和坟墓在美国中西部被发现(也见赛勒斯戈登教授,哥伦布之前,试金石出版社,1972)。
跨越大西洋一个奇特的东西是,据说有民族似与威尔斯人或古布列颠人有联系,但我们现有的记录并未显示有任何数量的R1b单倍群。这方面需要进一步的研究。各印第安部族的 脱氧核糖核酸,特别是那些在维吉尼亚和东北部者需被计量,因为据报导欧甘文字在那儿发现–因此它若发生脱氧核糖核酸应会揭露接触。对北美洲印第安人进行的人类Y染色体 检测至今只揭示了Q, Q3和C3。
我们已看到:
单倍群R有来自非洲至印度至澳洲的RxR1基本。
R1a包含大部分的斯拉夫人和印度/亚利安人,而单倍群R1b则包含大多数西欧人,两者为凯尔特和盎格鲁撒克逊/诺曼人。有大量的R1a扩展进入德国和挪威,比在西欧其他地区所发现的更多。我们在冰岛看到R1a, R1b和I开始与Q合并但只在这里(从爱斯基摩人在那里的掺入及传至挪威的一些家庭)。
若单倍群I是决定性的闪米特单倍群,那闪米特人散布在欧洲各地,与雅弗人混合在一起。巴斯克人、威尔斯人和爱尔兰人是较独特的单一单倍群人民(R1b),但即使是他们也有达到四个分开的单倍群,巴斯克人和威尔斯/康沃尔人只有两个单倍群。
可争论的是,闪和雅弗的单倍群都是F,而他们的儿子们是简单的变异没有差别,因此两组都有G至 J,K形成下一个分支而且是未出现在已知的闪米特J者。因此,K必包括所有的雅弗,但那也可争论G至I对两者开放。这些争论似未能承受以简单的人口基础作出审查。
进化论者争论说这些世系爬出了沼泽,他们的祖先活至十三万年前。J因而是闪米族世系,而他者都是独立的进化活动,但他们承认它全来自一个的祖先。他们争论说洪水是一个神活,任何认真看待圣经故事者对科学事实根本无知。
圣经作为灵感的神话,它必须以理性和真实的态度解释万民的根源。我们在这些看到的是,已知族群的脱氧核糖核酸与他们受确定圣经关系的其他族群相称。
另一个争论是,一切比我们所想像的发生的更快,这些变异在多个地点独立发生,而脱氧核糖核酸序列并不指示人民的根源。这前提在短期内与已知的检测和历史例子不符。
在这些所提出的建议与已知历史和圣经记录一致。它解释了遗传学的利用可在圣经记录内作出说明,并是利用延长时帧进化模型的可接受替代方法。
也可能的是,单倍群I是主要的闪米族单倍群,包括希伯来人并伸展至北部以色列人以及至以拦人。我们未在现代亚述人之中发现单倍群而只有G和J与L一起,一些单倍群I可能在古时发生。关于以色列人和亚述人会在末日复兴时手牵手一同走出北部,可能是因为他们皆离散在欧洲而且一些很可能是单倍群 I;但现代检测显示那些被确定为现代亚述人的族群中没有单倍群I。因此单倍群J是后期巴比伦被掳及犹太人和以实玛利人及以东人被吸纳入犹大之后,在阿拉伯国家中的中东发展。即使是在非常遍向公元前最早可能日期的进化模型中,J2科恩形式单倍群被也被接受为可能有二千一百年之久。因此,单倍群I很可能比它早数世纪并确定当时分散在欧洲各地的以色列十族。雅弗得福使他的后代能被置于闪的帐幕中而完成了预言,而闪所得的祝福归于雅弗,虽然雅弗在每个国民中的百分比较大。
可以确定的是,欧洲单倍群I和R1b以及R1a的纯粹程度使到欧洲的近乎每一个人,通过他们的母系或父系,实际上不可能不是亚伯拉罕、以撒和雅各的后代。因此,赫梯与以色列的联盟使他们成为一个民族,遍布广大地区有着各种语言实体。亚伯拉罕的确成为很多民族之父。