上帝的基督教会
编号
B9A
进化论和“非洲起源理论”
(第1.0版 1.0 20110116-20110116)
进化论声称现代人类在非洲发展,并于78,000年前从那里传播到世界各地。然而,圣经则有不同的说法。两种说法不可能同时正确。
上帝的基督教会
邮政信箱
369,
沃登
ACT 2606,
澳大利亚
电子邮箱:secretary@ccg.org
(版权所有
ã 2011 韦德·考克斯 )
在保持该文完整性,即没有改动和删除的情况下可以自由复制和传送。必须包括出版商的名称和地址以及版权声明。不得向接收分发副本者收取任何费用。在不侵犯版权的情况下,可以在评论性文章和刊物中作简要引用
本文可从以下万维网网址获取:
http://www.logon.org和http://www.ccg.org
进化论和“非洲起源理论”
本文将研究进化遗传学家有关“非洲起源”猜想的游戏,以及将人类起源扩展到克罗马农人,并将时间回溯到78,000年前(YBP)的说法。为此,这些学者将哈姆的血统与其他两个闪米特和雅弗的血统结合,并声称闪和雅弗都是含的后裔。这样做的效果是将挪亚的儿子们结合起来,并通过使用完全虚假伪造的单倍群树来延长血统,并通过使用完全虚假的突变树表来将日期延长到78,000
YBP,这是完全错误的。人们已经注意到了这种游戏,并且我们可以在诸如www.eupedia.com这样的网站上看到例证。
让我们在这里重建论点,以便我们可以通过检查正确的DYS序列来恰当地证明,这个论点是如何虚假的,时间框架比所宣称的要短得多,即使使用了他们虚假的突变序列时间表也是如此。该论点提出如下。原始的Y染色体祖先是:
Y
据称在非洲出现在78,000
YBP
第一个突变A在欧洲的列表中没有列出。这是因为Hg.
A与其他染色体没有任何联系,并且Hg.
B
专属于非洲和非裔美国人。
请参阅附录A
这个序列的不可能性
通过查看下面的Y染色体家族谱,我们可以看到这棵树被分解成了绝对禁止形成任何这种推测树的谱系。
首先,我们必须从挪亚的祖先根源和含单倍群的主要分支的基本结构开始。Hg.
A是第一个从主要祖先系分化出来的。Hg.
A突变为M91和P 97?。它没有保留其他线路中发现的任何基础联系。
祖先的主线是从挪亚到雅弗、含和谢闪。谱系为SRY0831、1
M41 2、M139、M168、P9。
其他的含谱系突变为M60、M181和P85?。在HgB.的主要的Hg.谱系A和B或多或少地被限制在非洲和非裔美国人中。
其他来自M168和P9中心挪亚谱系的含谱系突变为库实特人血统的RPS4Y M216的Hg. C,以及M145、M213形成的Hg.
DE,这是YAP的分界线,且涵盖了Hgs.D血统的西藏人和Hgs.E的所有迦南人和北非血统。
因此,显然,主要的非洲谱系A和B与其他非非洲谱系没有任何关联,除了Hg. E进入并传播到北非和中东,然后再传播到欧洲这一事实。
闪和雅弗的所有谱系都以突变P14
M89和M213中的Hg.
F谱系分布在世界各地(根据2005年FTDNA家族谱图)。经过检查,发现M213在DE和F等中是常见的,但对Hg.
C不常见。因此可以推断,M213在C组中突变为M216,并插入了RPS4Y。而在Hg.
DE中,M145在M213之前被插入,在Hg.
F和随后的谱系中,P14和M89在M213之前被插入。因此可以断言,M213是闪、含和雅弗的原始挪亚血统的最后一部分。无论他们声称这种突变需要多长时间还是多短时间,这都是科学测量的Y染色体树的正确谱系。
因此,Hg. C、D、E和F以及所有随后的G、H、IJ和K群体不可能来自A或B,也不可能像“非洲起源”论点中所声称的那样来自C。
圣经记载和血统的突变组表明,主要的突变发生在五次自发突变中,远离了主要的挪亚血统。然后,它们陷入随后的突变中,形成了其他部落/民族群体。提出相反观点似乎是故意欺诈行为。
从他们自己的图表中可以看出,这些联系和时间框架都是虚假的,图表上42,000
YBP之前的所有事件也都是虚假的。
所提出的其他突变的分解同样是完全的猜测,也是假的。因此,这些事件甚至不能以42000
YBP的日期为基础。
如果忽略错误延长的图表,并将圣经模型放入透视中,并适当讨论和考虑现代背景辐射和线粒体DNA对人类基因组的影响,那么从公元前2200年至今各种突变中血统的分解实际上都可能发生。进化论是一个宗教神话,必须得到适当的解释和驳斥。然而,为了维护这种宗教神话,科学必须被篡改,关键要素必须被忽视或歪曲。
这种失实的陈述经常发生。最近的一个案例是在俄罗斯和以色列发现的类人生物化石,这可以追溯到2003年发现的所谓弗洛雷斯的霍比特人。
在俄罗斯西伯利亚洞穴中发现的指骨年代为30000 YBP,但与往常一样,年代测定的基础没有公开讨论。据报道,它有一些异常完好的DNA。
科学家们提到最近在俄罗斯发现的东西时,又提及了“弗洛雷斯的霍比特人”,称其有超过12,000年的历史,并有人甚至认为它是30,000年前的一个古老的、非现代化的人类物种。这尽管科学家已经调查过弗洛雷斯的发现,并且从其Y染色体中证实,它是一种由于在弗洛雷斯的饮食导致退化并缩小了体积和容量的现代人类。这实际上是一个如何不去生活以及如何丧失人类社会能力的例子。
然而,这些科学家却利用了这些数据,他们要么知道并故意滥用这些数据,要么连打开电视都不愿意,他们肯定从未研究过有关这个问题的最新调查。进化论的高级祭司使得中世纪罗马天主教修士看起来更具进步性。
2010年12月发表在《自然》杂志上的发现证实,这个标本来自一名既不是“现代人类”也不是尼安德特人的年轻女孩。相反,据报道,她属于人类家族树上的一个独立的、现已灭绝的分支,科学家们以化石发现地西伯利亚南部的丹尼索瓦洞穴命名为丹尼索瓦人。
此外,据报道,从洞穴中发现的一颗臼齿被认为来自丹尼索瓦人。据报道,它看起来与现代人类和尼安德特人的牙齿不同,而且据称更像直立人等更古老的人类祖先的牙齿。。
基于他们对DNA结构的推测,科学家们接着断言,这些发现改变了人类进化的故事。然后他们继续断言这些发现表明,尼安德特人有一个亚洲的姐妹群体,这个群体在他们自己的进化道路上分裂出去后灭绝。尼安德特人与人类有着完全不同且绝对陌生的DNA系统,这一事实被忽视了,随后这一发现被应用于这样一个假设:这些史前人类与尼安德特人等史前人类有某种关系,然而我们知道他们并没有这种关系。
大多数科学家认为尼安德特人的祖先于30万至40万年前离开非洲,在欧洲和亚洲建立自己的家园。科学家们随后声称,同时,现代人类的直系祖先智人留在非洲,并在那里进化。
据称,他们约在70,000到80,000年前离开非洲,并曾一度与即将灭绝的尼安德特人共存。
现在,科学家们认为丹尼索瓦人在最初的30万到40万年前的大迁徙时期与尼安德特人分化,并向东扩张。
然而,与所有的DNA证据完全相反,科学家们断言:“就像尼安德特人一样,他们似乎与现代人类交配过。”我们与黑猩猩杂交的机率比与尼安德特人的机率更大。至少黑猩猩有一个八链系统;尼安德特人没有。
据称,这些证据意外地出现在现代美拉尼西亚太平洋岛民的DNA中,其中包括了来自丹尼索瓦人的基因序列。我们可能会问,这是怎么发生的呢?美拉尼西亚的YDNA结构是基于Hg.
K序列的,我们知道这是F的直系后代,并且存在于上面的序列中。这个K序列中的哪个神秘部分是从东方的直立人那里衍生出来的?也许Hg.
K亚当时代的某个人类与一个史前人类的遗迹杂交了;尽管这种假设没有任何证据支持。
研究表明,至少在一些美拉尼西亚人的遗传密码中,来自丹尼索瓦人的遗传物质约占4%至6%。是这样吗?涉及哪些DYS,以及这种断言的依据是什么?美拉尼西亚人是猩猩吗?涉及哪些mtDNA?我们知道,所有人类的mtDNA源自mtDNA
Hg. L。他们对这种断言有什么可能的依据?他们之所以不公开披露这些说法的依据,是因为一旦暴露在新鲜空气和阳光的有效防腐剂下,这些说法总是错误的。
本周在以色列发现的八颗牙齿也据称是现代人类的祖先。他们声称这是40万年前的。事实是,以色列是尼安德特人的发现地,它们都经过了测试,与现代人类毫无关系。
从以色列的发现中,他们能够证明他们烹饪并食用植物和蔬菜。据报道,美国的研究人员在他们的牙齿中发现了煮熟的植物材料。这项研究首次证实,尼安德特人的饮食不仅限于肉类,而且比以前想象的更加复杂。该研究已经发表在《美国国家科学院院刊》上。
与大多数这类说法一样,迄今为止的证据都是间接的。它们明显是食人族,对骨骼进行的化学分析表明他们很少或根本不吃蔬菜。一些人认为,这些人类对肉类的依赖是导致这些类人动物灭绝的原因之一,因为大型动物如猛犸象减少了。
然而,科学家们对全球范围内的尼安德特人遗骸进行了新的分析,发现了直接证据与化学研究相矛盾。研究人员在他们的牙齿中发现了化石化的植物颗粒,其中一些是煮熟的。
尽管以前在尼安德特人的遗址和一些洞穴中发现过花粉粒,但但直到现在才有明确的证据表明这些人实际上确实吃过植物性食物。
“我们以前在尼安德特人的遗址上发现过花粉颗粒,但你永远不知道它们是被吃掉还是被睡觉时放在下面的。”
“但在这里,我们有一个案例,其中植物的一小部分在口腔中,所以我们知道尼安德特人正在食用这种食物”(乔治·华盛顿大学的艾莉森·布鲁克斯教授对BBC表示)。
测试结果错误的原因,使得大部分发现都不准确,这可以归结为假设,这些假设来源于分析人员自身的先入为主的观念。
布鲁克斯教授表示,这些测试是在测量蛋白质水平,研究人员假设这些蛋白质来自于肉类。
“我们过去倾向于认为,如果饮食中的蛋白质含量非常高,那一定来自于肉类。但是……有可能其中一部分蛋白质来自于植物,“她说。
据称,这项研究是最新的,它表明,“尼安德特人远非野蛮的野人,他们与我们的相似程度比我们以前想象的更高。”
尽管DNA中有绝对证据毫无疑问地表明我们与他们或他们与我们无关,但这种寻找缺失环节并将前亚当时代人类与亚当时代人类联系起来的动机却不断被忽视和压制,以维持进化宗教的神话。
现代科学家身负进化理论的包袱,与他之前的三位一体宗教体系中的前辈一样受到束缚。毫无疑问,进化是一种基于神话和猜测的宗教。
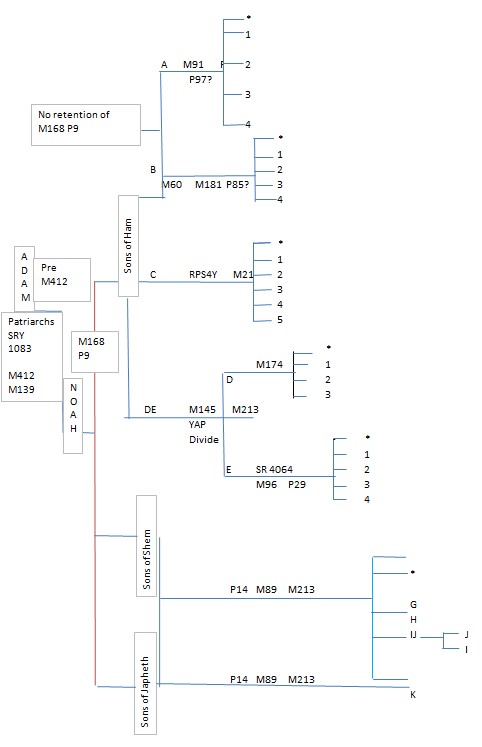
我们已经讨论了上面的各个国家的族系,并展示了现代人类78,000年前(YBP)不可能来自非洲的情况。下面的附录B显示了从YDNA家谱树直至Haplogroup
F(单倍群F)。
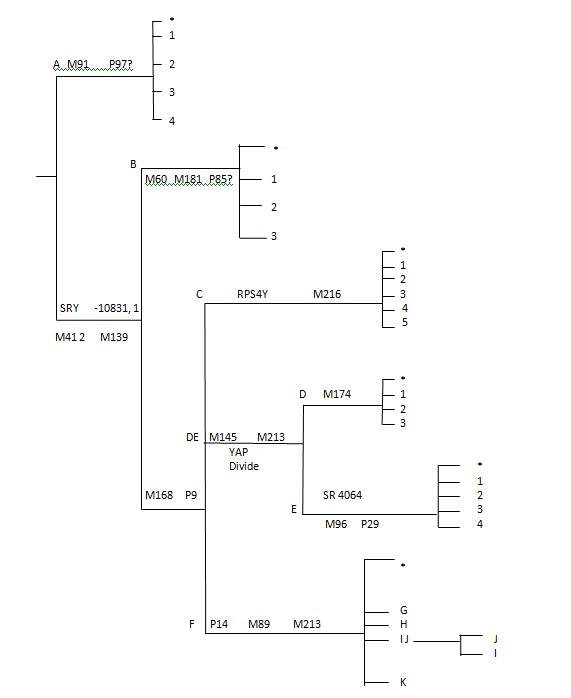
我们现在将重新排列YDNA树,以便更容易或更清楚地显示诺亚和他的儿子闪、含和雅弗的YDNA谱系,以及突变是如何从该谱系发生的,以及它们是如何在闪和雅弗之子中进一步突变的
挪亚在他的时代是纯洁的(创6:9)。RSV版本将其翻译为:“他在当时的世代是个完全人”,但通常人们认为这段文字是指他的遗传结构。
从亚当开始的YDNA序列基本上是M412、M139的结构,它构成了现代人类YDNA结构的核心。挪亚将M168和P9的根本能力传给了他的每一个儿子。含将核心能力或能导致Hgs.
A、B、C和DE突变的遗传先决条件传给了他的每一个儿子。这些突变的性质是这样的,使我们不得不从这些突变和随后发生的mtDNA突变的分布中得出一些必然的结论。我们可以从这些突变和分布的逻辑中得出一些惊人的结论。
我们必须做出的第一个逻辑推断是方舟上有多少人。圣经说有八个人。这个问题的答案引发了一些严重问题,关于这些文本所指的是什么。当提到这八个人时,我们是指四男四女吗?这是多年来圣经学生的普遍假设。他们被认为是一夫一妻的事实也被认为是理所当然的。有许多圣经和科学上反对这种观点。
圣经还说,第一次收获葡萄酒,一定是在洪水后的三年内,诺亚喝醉了。他随后受到了虐待。再加上迦南活着并年龄足够大,可以对他的祖父挪亚的行为负责,他受到了严厉的咒诅和惩罚。面对如此严厉的惩罚,这是一个正义和法律的问题,迦南必须明显是个成年人才能接受这样的审判。他是含的儿子中最年轻的一个。因此,他们一定都在方舟上。再加上对单倍群突变的其他考虑,很可能有八个男人,其中包括挪亚和他的三个儿子以及其他四个是含的儿子。这就引发了关于母亲的问题。他们是同一个女人的孩子,还是不同女人的孩子?我们将从基因的突变和圣经故事中看到,尽管它们可能具有略微相似的mtDNA结构,但它们仍然是不同的。
我们现在可以确定,原始女性mtDNA属于单倍群
L。我们还可以确定,原始Hg.
L现在仅限于非洲;因此,“走出非洲”假说的起源也就清楚了。几乎可以肯定,夏娃的血统是深色皮肤,或者至少她具有黑色素和生理能力发展成现在的现代非洲女性。她属于单倍群
L,就像原始的mtDNA群一样。DNA科学认为突变速率非常缓慢,事实上远远慢于它们的实际速度。此外,科学曾假设mtDNA不影响人类基因组,不参与基因突变。这两个假设现在都被证明完全错误。这些变化的基础在论文《民族的遗传起源》(
第265期)中有所讨论。
确实,正是这篇论文确定了Hgs.
I和J作为最初的希伯来人Haplogroup
IJ,后来分裂为两个群体的逻辑必然性。科学家们最初拒绝了这一假设,但他们最近在结构的S2和S22链接处发现了联系;尽管他们随后宣布Hg. I是原始闪米特族的,并将其追溯到30,000年前。
背景辐射量影响人类DNA的突变速率。已经证明,人类mtDNA在高背景辐射区域的突变速率为每代最多一次突变,正如我们在上述论文中提到的喀拉拉邦实验中看到的那样。此外,已经证明mtDNA会导致整个人类基因组的突变。因此,背景辐射会使mtDNA的突变速率大幅增加,就像它对YDNA的影响一样。这些mtDNA的突变独立于背景辐射,进一步增加了突变速率。
因此我们可以看到,当两种突变效应结合在一起时,由于mtDNA的进一步突变而导致的突变速率必然以加速的顺序增加。因此,突变速率可能比科学假设的要多得多,以便证明他们的进化模型。整个论证结构都建立在已知的错误前提上,并必须在适当调整之前被逻辑上驳回。
我们还可以通过已知的DNA结果证明,原始的DNA与已知的mtDNA基因组相关联并一起传播。例如,早期发现的澳大利亚原住民发现在八波有三个YDNA
单倍群和三个mtDNA
单倍群。这些是YDNA Hg.
C4(突变为C4a和C4b)、Hg.
K和Rx
R1basic。女性mtDNA
单倍群与男性群几乎完全匹配。从这项研究中我们可以看到,闪族C4组与女性Hg.
N Basic相匹配,这是从L3突变而来的第一个突变。
因此,我们可以推断L,作为原始物种,看到方舟上的分裂是L的突变,也许还有L1或L2(尽管L2可能是L的后续突变)和L3(突变为N,然后是M)。L3最早的分裂到N
Basic必须发生在C4移入澳大利亚之前。这些联系必须是库希特族Hg.
C最早从中东进入亚洲,然后进入澳大利亚的移动。因此我们可以推断,Hg.
N基因的突变必须发生在L3最早的时期,要么是洪水之前,要么是洪水之后。有可能Hg.
N基因来自挪亚儿子的至少一位妻子,并且似乎也在方舟上。由于它是突变的基础,可以推测它至少在两个,可能是四个妻子身上。另一个推断可能是,由于N基因在非洲并不是主要因素,因此含的儿子,他们是YDNA
单倍群 A和B,并没有与该突变的女性结婚,而是保留了来自基础L1和L2突变的妻子。
为了形成L2,必须在方舟上存在L,或者L2在之前就形成了,并且L1、L2和L3的所有三种突变都存在。由于从L没有进一步的突变,原始的L必须在此之前见过突变,而原始的L可能已经不存在了。因此方舟上必须有mtDNA
Hgs. L1(或许是L2)、L3,而且可能有N基因型。由于mtDNA
Hg. M是从其他单倍群中突变而来,Hg. R也是从Hg. N中突变而来,所以它们不一定需要在方舟上。
很明显,L存在一个瓶颈情况,并且从L、L1和L2结构或者从那里没有进一步的突变,所有后续的突变都来自mtDNA Hgs. N和M,随后又来自于N作为Hg.
R超群和随后的群体。这导致了关于N和M等的突变是在非洲之外发生的说法。圣经的解释是,这种情况发生在中东,而含的儿子被赋予非洲作为他们的继承地,并在洪水之后和分散之后移居到那里。事实上,发现YDNA
Hgs. C、D和一些E远离非洲并遍布世界各地并不符合“非洲起源”的理论逻辑。
事实上,它支持相反的理论。非洲板块实际上包括约旦断裂带西侧,一直延伸到黎巴嫩的海岸。因此我们可以得出结论,其中一些人是“在非洲”。然而,古实安全地或胡斯在幼发拉底河以北。事实上,从圣经的角度来看,伊甸园的土地一直延伸到埃及和尼罗河以及地中海沿岸。这可能是弥散的逻辑候选地点,而这也正是圣经所记载的。
很可能YDNA Hgs.
A和B的妻子是L1和L2,因为这些群体一起进入非洲并且留在那里。其余的妻子则是mtDNA
L3和N基因型。L3除了在已知的后期迁徙中之外,在非洲以外的地方没有发现。因此N似乎是非洲以外的主要mtDNA来源。M也可能在那里,但N似乎覆盖了其他基因型并发生了突变。很可能mtDNA
Hg. N是挪亚的儿子中最多的mtDNA之一。含的另外两个儿子很可能也娶了Hg.
N,正如闪和雅弗所做的那样。这可以更容易地解释基因分布和随后的突变。
N基因型与C4的存在表明mtDNA
Hg. N的女儿们非常早地就嫁入了库希特族,或者最初就在那里。还要注意的是,在YDNA
Hgs. A和B中没有保留M168 P9。这要么是因为他们在巴别塔干预之前就进入了非洲并且最初没有它,要么是因为他们在那里的病毒/辐射活动中失去了它。Hg. A从早期的序列中发生了突变,或者看起来是这样。
我们可以看到,我们在DNA科学中所知道的任何事情都不支持圣经的结构,事实上它有助于更有效地解释圣经中的问题。与圣经相冲突的事物,在检查时表明长期的推测是错误的,并且试图延长时间框架的尝试是基于欺诈性的伪科学。请参阅附录C中关于分配给挪亚子孙的YDNA系统的血统支系的图表。
q
附录A
进化的“非洲起源”模型
|
然后,该谱系据称发生了突变到 BT |
74,000年前 |
|
然后,该序列据称进展到了 CT |
70,000年前 |
|
这一突变据称与第一次从非洲迁移的时间相符 |
|
|
然后,该谱系据称分成了两个分支: |
|
|
DE |
大约65,000年前 |
|
另一个分支CF(两者均源自Hg. C序列)发生了 |
大约58,000年前 |
|
DE据称从非洲分裂成Hg. D和E,形成Hg. E |
大约54,000年前 |
|
CF继续突变成Hg. F |
大约48,000年前 |
|
Hg. F然后分裂成: |
|
|
Hg. IJK |
大约45,000年前 |
|
以及Hg. G |
大约32,000年前 |
|
(Hg. G突变形成G2a 12,000年前,见下文) |
|
|
据称Hg. IJK分裂形成: |
|
|
Hg. K和 |
大约36,000年前 |
|
Hg. IJ |
大约32,000年前 |
|
Hg. IJ据称突变形成J,和 |
大约29,000年前 |
|
Hg. I
在 |
大约24,000年前 |
|
据称Hg. I形成于德国地区,这是不可能的。我们将在稍后查验这个虚构的内容。 |
|
|
据称Hg. K突变成NOP |
大约36,000年前 |
|
Hg. K还据称突变形成Hg. R从非洲出发 |
大约30,000年前 |
|
Hg. NOP据称突变形成Hgs. NO和P |
大约29,000年前 |
|
Hg. NO据称突变形成N |
大约22,000年前 |
|
P然后据称突变形成Hg. Q |
大约24,000年前 |
|
以及Q1a |
大约20,000年前 |
|
据称Hg. I形成于22,000年之前,然后是Hg. I2 |
大约17,000年前 |
|
G2a据称形成于12,000年前,然后G2a3据称形成 |
8,000年前 |
附录B
典型YDNA家谱树
附录C
应用于圣经结构的YDNA树